Campylopus, a modern and successful genus!?
|
|
|
- Briana Fitzgerald
- 10 years ago
- Views:
Transcription
1 91 Tropical Bryology 2: , 1990 Campylopus, a modern and successful genus!? Jan-Peter Frahm Universität Duisburg, Fachbereich 6, Botanik, Postfach , 4100 Duisburg, W-Germany Abstract. Campylopus with nearly 200 revised species is one of the largest genera of mosses in the world and has extremely broad geographical, altitudinal and ecological ranges. Factors to be considered for the rich speciation are 1) the enormous anatomical plasticity of the structure of the costa, 2) the ability for vegetative propagation utilizing different methods, and 3) the special twist mechanism of the cygneous setae. Phytogeographical interpretations of present ranges of species of Campylopus lead to the conclusion that this genus is of Gondwanalandic origin. Most of the species seem to be cool temperate in origin. They have adapted to dry habitats in the Mesozoic and invaded the tropical mountains during the Tertiary, accompanied by rich speciation. This is supported by the physiology of these species. According to preliminary gas exchange measurements, tropical montane species do not differ from temperate species, and are not able to stand climatic conditions of the tropical lowland rainforests; a relatively young habitat for bryophytes. A cladistic analysis of the infrageneric categories of Campylopus using phytogeographical evaluations of apomorphic character states shows that the section Homalocarpus seems to be the most primitive and the subgenera Campylopidulum and Thysanomitrion the most derived. 1. Introduction The somewhat provocative title goes back to Schuster (1984), who (as a hepaticologist) interpreted some distribution maps of Campylopus and called this genus modern and successful. So it would be interesting to prove whether a moss genus such as Campylopus is really modern and successful. In total, more than 1000 species of Campylopus have been described. Twenty years ago, Index Muscorum (Wijk et al. 1959) listed more than 700 legitimate species. Since that time, the number has been reduced by worldwide revisions to less than 200. This is a common effect in bryophytes, especially in tropical ones, where rates of reduction owing to worldwide revisions, are about 60-70%. This indicates that there are not species of bryophytes in the world as given in textbooks, but probably no more than (4.000 species of hepatics and species of mosses). There are only a few genera of mosses with numbers of species comparable to Campylopus, but these have not yet been revised critically on a worldwide basis, and therefore Campylopus may be one of the largest genera of mosses in the world. Based on an estimated total of about 6000 species of mosses, i of every 30. mosses (or 3.3% of all mosses) belong to the genus Campylopus. The genus Campylopus has a wide latitudinal range between 65 0 N (Alaska) and and 65 0 S (South Sandwich Islands) and an enormous altitudinal range between sealevel and more than 4500 m elevation. The species occur on a variety of different substrates such as nutrient poor sand, humic soil, peat, rocks, rotten and living wood,


2

3 Gametophyte Costa The most conspicuous vegetative character of the genus Campylopus is the broad costa with extreme diversity of internal differentiation. There is no comparable range of variation in the structure of the costa in any other moss genus. All the different expressions between types with two stereid bands and a median deuter band (as commonly found in the Dicranaceae and also other families) and costae with no stereids at all can be explained by a few cell divisions (fig. 1). Thus the mechanism behind this broad variety of structures is relatively simple. What are the advantages of these different costal structures? a. mechanical stability. Stereids with firm thickened walls, provide mechanical support either in layers on the adaxial surface or in bundles in the abaxial part of the costa. They are supposed to be good protection against shrinking caused by desiccation and are therefore are found frequently in dry adapted species. In species pairs (often sympatric species differing mainly in one character of the costa and the habitat), the species of the wetter habitat has ventral hyalocysts, whereas that of the drier habitat has ventral stereids. One of many examples is Campylopus comosus (Reinw. & Hornsch.) Bosch & Lac. growing on soil, with small ventral hyalocysts and C. flagelliferus (C. Müll.) Jaeg. growing in the same range but epiphytic with ventral stereids (Frahm 1985, Frahm et al. 1985) b. Photosynthesis. Assimilation is carried out by a median band (the so called deuter cells) and a dorsal band of chlorocysts. Since the width of the costa is usually more than half of the leaf width, and the basal laminal cells are often nonchlorophyllose, the costa is the main source for photosynthesis. c. Water uptake and storage. Water is easily taken up and stored by hyalocysts, especially the large ventral ones. This seems to function only in wet habitats, since the cell walls of large hyalocysts are very delicate and can probably not stand desiccation. In species pairs the species of wet habitats have larger hyalocysts than species in drier habitats, as exemplified by Campylopus shawii Wils. growing in swamps and C. setifolius Wils. growing on wet cliffs (Frahm 1985). It seems, however, to be paradox that species of wet habitats should depend on storing water, where there is no dry daily or annual season. Thus it may be that the function of the hyalocysts (as well in the Leucobryaceae) have been misinterpretated. As Robinson (1985) has observed, the hyalocysts in the Leucobryaceae often show air bubbles, and the major function may not be water storage but gas exchange. This hypothesis is supported by the location of the chlorocysts, which are situated in the middle of the leaf without any direct contact to the atmosphere. Another method of water storage is between the dorsal lamellae of the costa. In species pairs, the species of drier habitats have the longer lamellae, for example Campylopus pilifer Brid. and C. introflexus (Hedw.) Brid. (Frahm 1985). d. Water conducting is probably possible not only from cell to cell or through the pectine lamellae of the walls but also more efficiently through bundles of stereids. In some species the alar cells (functioning for water uptake from the water conducted along the stem) bulge into the costa, indicating a conducting pathway from the alar cells to the costa. e. Gas exchange combined with photosynthesis. Effective gas exchange requires a large surface. Lamellae (being chlorocysts) seem to function not only for water storage but also for gas exchange as shown by Campylopus pilifer Brid. var. lamellatus (Mont.) Gradstein & Sipman, which is not found in arid environments like C. pilifer s.str. but in rainforests. In this kind of habitat, water storage is not required, but gas exchange in high humidity may be a problem.
. Thus the mechanism behind this broad variety of structures is relatively simple. What are the advantages of these different costal structures? a. mechanical stability.")
4 94 Thus the plasticity of different structures of the costa allows adaptations to a broad variety of habitats. The facility to modify such structures has promoted the broad ecological and geographical ranges and enhanced the high number of species Vegetative propagation Compared with other genera of mosses, vegetative propagation is relatively important in Campylopus. In this genus, all different kinds of vegetative propagation from microphyllous branches and deciduous leaves to special brood leaves, broken leaves and stem tips are found. If 40% of all species are known only sterile, the importance of vegetative propagation is evident. The vegetative propagation allows a species to survive in a habitat as a unisexual clone. It also allows clones to survive long unfavourate climatic conditions with no sexual reproduction. At least it allows propagation over short distances until clones of different sexes meet and allow fertilization. Apparently most of the species (if not all) are able to vegetative propagate by more than one kind of vegetative propagation Perichaetia Differentiated bud-like perichaetia function for splash cup dispersal of spermatozoids and also as protection for the young sporophyte during the stage of development when the seta is curved into the perichaetial leaves. Such perichaetia are found in all infrageneric categories of Campylopus. 2.2 Sporophyte The major characteristic of this genus is the seta which is curved sigmoid in the middle and twisted in the upper and lower part and uncoils when moistened. This character which is also present in genera such as Microcampylopus, Campylopodium, Dicranodontium or Campylopodiella (but not in other closely related genera such as Atractylocarpus or Pilopogon). The sigmoid curve in the seta is caused by its development (Frey & Frahm 1987 figs. 1-7). When the seta has reached half of its final length, it curves down, the capsule developes, often leaves the calyptra between the perichaetial leaves and curves up again. In this way the young sporophyte developes protected in the perichaetial leaves which is an advantage, especially in exposed or dry habitats. The twist mechanism of the seta causes wide movements of the capsule in a moistened state at the same time as when the peristome is open. The function of these circular movements of the capsule is not clear. It may enhance spore dispersal to all directions, but this is a relatively small advantage compared with the anatomical structures necessary for these movements. Spore dispersal in wet weather conditions also prohibits long distance dispersal. However, since Campylopus as many other genera of Dicranaceae are dioicous, male and female spores need to land near to one another in order to establish new male and female clones which are situated close enough to allow fertilisation. The mechanism for these uncoiling movements have not been known yet but could be explained by a study of the ultrastructure of the setae (Frey & Frahm 1987). The epidermal cell layer of the setae possesses a three layered wall with asymmetric lumens. SEM and TEM studies revealed a distinct lamination in the second layer and small pores in the outer, asymmetrically thickened epidermal cell layer of the seta, and a firm inner teriary cell wall. In dry state the setae are coiled by spirally arranged microfibrils in the secondary cell wall. Water (or water vapour) is taken up through the outer cell wall, stored between the microfibrils of the secondary wall, causing elongation of the setae and uncoiling movements. Opposite twist directions in the upper and

5 95 lower part of the setae prevent the setae from being twisted off. 3. Distribution and Speciation Campylopus is an ideal genus for phytogeographic interpretations because of its wide distribution, broad ecological range, wide altitudinal range and high number of species. Insofar it is equivalent to several other genera or even one or more smaller families. The worldwide distribution of the genus is uneven on both sides of the equator. There are 14 species in the subantarctic but only 1 species in the subarctic (Frahm 1988). In the tropics, the species of Campylopus are mostly confined to the montane belt. There are only a few species occurring in the lowlands, mainly in savannahs and not in lowland rainforests. The species in the temperate belt of the northern hemisphere are (1) either mainly tropical montane species occurring as remnants of the Tropical Tertiary flora in oceanic regions (cf. many species of North America and Europe, Frahm 1980, 1984), or (2) are of subantarctic origin and probably introduced into the northern hemisphere (cf. Campylopus introflexus (Hedw.) Brid. in North America and Europe, Frahm 1980, or C. pyriformis (Schultz) Brid. in Europe, cf. Corley & Frahm 1982), or (3) are endemics related to sister species of tropical areas (cf. Campylopus setifolius Wils. derived from C. shawii Wils., Frahm 1985a). This leads to the suspicion that the genus may have its origin on the southern coast of Gondwanaland and has spread to the tropics. This pathway is illustrated by several subantarctic species, which have closely related species in the tropics, sometimes 3 such species derived from the same (still present) ancestor in South America, Africa and Australia (Frahm 1988), representing a species group with a species mosaic of several small, mostly allopatric but also sympatric species. There are apparently several ways of speciation: 1. adaptations to dry habitats. For example, Campylopus introflexus (Hedw.) Brid. can be regarded as the ancestor of C. pilifer Brid., adapting to dry habitats by longer costal lamellae for storing water. Both species show a gondwanalandic range: C. introflexus along the south coast (today circumsubantarctic), C. pilifer in the interior (today from South America to India). By the same way, C. julaceus Jaeg. (with one subspecies in SE-Brazil and one in E-Africa) and C. aemulans (Hampe) Jaeg. (in SE-Brazil) can be derived. Campylopus surinamensis C. Muell., a neotropic savannah species, is derived from the subantarctic C. vesticaulis Mitt. 2. Adaptations to alpine habitats. There seem to be two pathways to alpine habitats, one from the subantarctic, especially through the Andes, which is well known for many other tropical alpine bryophytes, and another one from montane species. There are several pairs of species or subspecies, one of them confined to montane, the other to alpine habitats. Examples are C. albidovirens Herz. in subalpine forests of the Andes and C. pittieri Wils. in the Paramos; C. flexuosus (Hedw.) Brid. ssp. flexuosus in the montane regions and ssp. incacorralis (Herz.) J.-P. Frahm in the subalpine regions of the Neotropics and Africa, C. cuspidatus (Hornsch.) Mitt. var. cuspidatus in montane rainforests of the Neotropics and var. dicnemioides (C. Müll.) J.-P. Frahm in the alpine. Other examples are C. amboroensis Thér. in the high Andes and C. johannis-meyeri (C. Müll.) Kindb. in the afroalpine mountains, both derived from C. pilifer Brid. The origin of the alpine taxa may be explained as ecological adaptations, which may have been supported by extensions of the alpine belt during the fluctuations of vegetation belts in the Quaternary. Interestingly, one new habitat like the páramo has been colonized by different taxa from different relationships within the genus, resulting in very similar
. In the tropics, the species of Campylopus are mostly confined to the montane belt.")
6 96 adaptations. For instance, Campylopus pittieri Williams and C. cavifolius Mitt. both grow often intermixed and can hardly be distinguished in the field. Only transverse sections of the costa show that these two species belong to different systematic groups. There are similar analogous developments in Dicranum. Species adapted to alpine habitats show the same growth form (compact cushions, orthophyllous appressed leaves, subtubular leaf apices). They show, however, significant differences in the structure of the costa and the shape of the capsules (for example D. groenlandicum Brid. with erect capsules and two stereid bands, D. brevifolium (Lindb.) Lindb. with curved capsules and two stereid bands, and D. muehlenbeckii B.S.G. with curved capsules and two stereid bands and two epidermal layers). This demonstrates that species from different evolutionary lines have adapted to similar habitats. The only species of Campylopus occurring in the subarctic and alpine regions of the northern hemisphere, C. schimperi Milde, surprisingly shows the closest affinities to subantarctic species. It is, however, not found in the Andes of South America or the African mountains. This may be an example for a species which has drifted from the southern hemisphere to Asia via the Indian subcontinent and has spread through the northern hemisphere. This pathway has been pointed out by Schuster (1972, 1976) for hepatics, Smith (1972) for Polytrichaceae, and Raven & Axelrod (1974) for Angiosperms. 3. Adaptations to different habitats: This concerns sympatric species which occurin different habitats, especially epiphytic species, that are derived from terrestrial ones. Examples are C. comosus (Reinw. & Hornsch.) Bosch & Lac. and C. flagelliferus (C. Müll.) Jaeg. in SE-Asia, or C. lamellinervis (C.Müll.) Mitt. and C. huallagensis Broth. in South America, the latter species being epiphytes. By the same differentiation, Campylopus setifolius Wils. seems to have split from C. shawii Wils. in the British Isles by changing habitats from swamps to wet rocks. 4. Physiology The cool-temperate origin of the species of Campylopus as that of many bryophytes (Schuster 1984) and probably most tropical montane mosses is supported by their physiology. These species lack the ability to descend to the tropical lowlands, as measurements of the gas exchange have shown (Frahm 1987, 1987a), due to high rates of respiration. Physiologically seen tropical montane bryophytes react as do cool-temperate species. Only a few species were able to adapt to the humid tropical lowlands, probably the youngest in evolution. In Campylopus, there are no lowland rainforest species, and only a few lowland savannah ones. The majority are montane in distribution. Tests in climate chambers show that species such as Campylopus flexuosus cannot stand the ecological conditions of tropical lowland forests characterized by high temperatures and low light intensities. 5. Cladistics A cladistic analysis could give an idea, which species posses the most primitive characters and which have the most advanced features in this genus. This could also provide a hypothetical background of the origin of this genus. It requires, however, the evaluation of primitive and derived characters. This is a major problem in this genus because of the enormous variation of characters. For example, the capsules may be asymmetric or symmetric, the calyptras ciliate or nonciliate, the basal laminal cells hyaline and thin-walled or incrassate and pitted. There is some support for the evaluation of morphological character states by the interpretation of ecological, phytogeographical phytochemical, developmental and palaeobotanical facts.


7
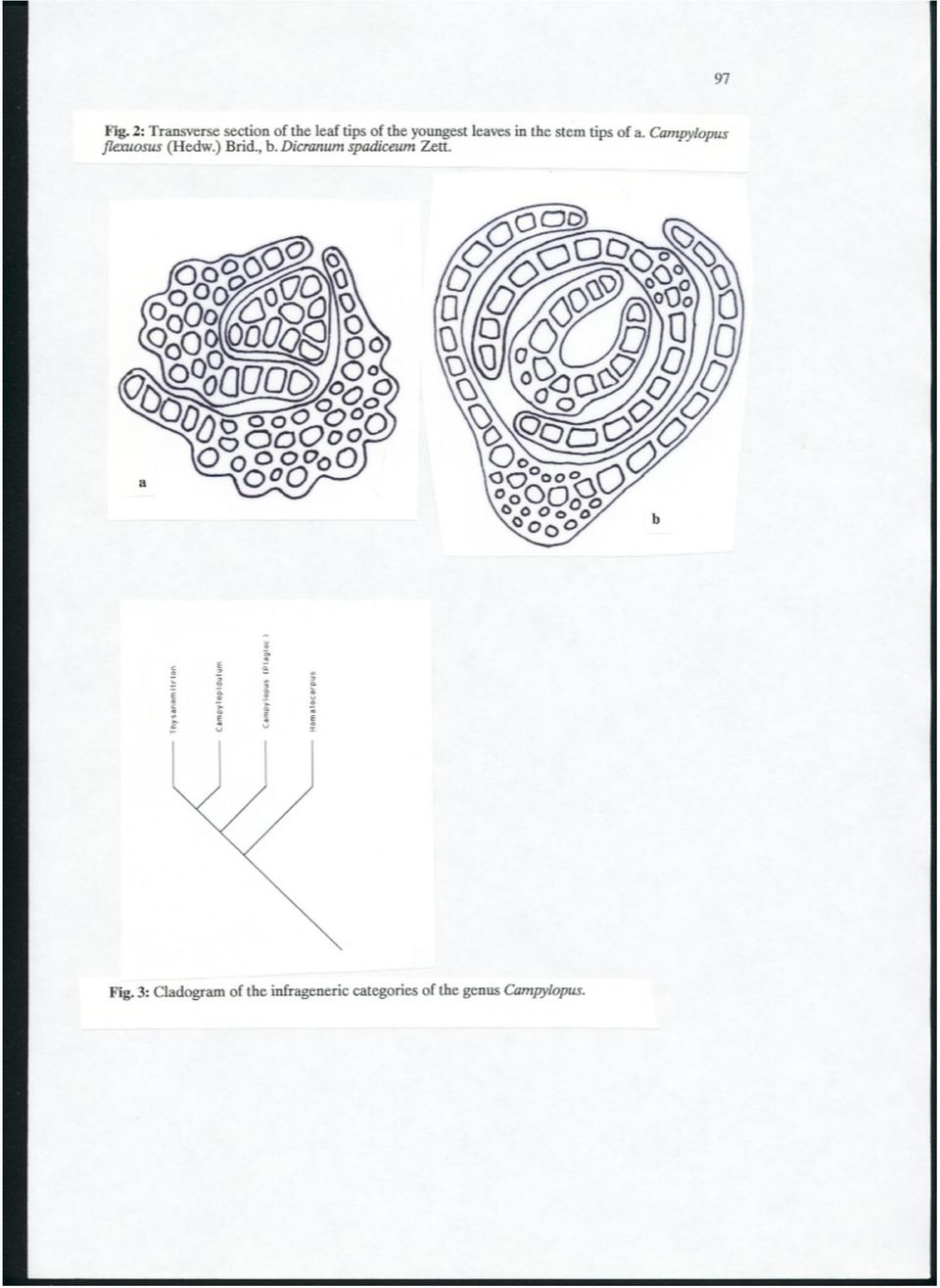
8 98 It is in ecological evidence that epiphytic species (found only rarely in this genus) show apomorphous character states such as incrassate laminal cells and stereids in transverse section of the costa. These are the same adaptions as found in dry adapted soil species. There is phytogeographical evidence that the subgenus Thysanomitrion has most of its representatives in SE Asia but only one in the subantarctic, which is the probable ancestor and its character states can be regarded as plesiomorphous. There are only a few phytochemical data present for this genus (Frahm 1983). These data show that certain species in the section Homalocarpus have no flavonoids whereas the highest diversity is found in the subgenus Thysanomitrion. A major problem is in the interpretation of the costal anatomy. Are transverse sections without stereids and large hyalocysts or those with ventral and dorsal stereids pleisiomorphe? Within the Dicranaceae, the most common type of costa is that with ventral and dorsal stereid bands. Therefore for genera such as Dicranum, species with tendencies to larger cells instead of stereids are regarded as derived (Peterson 1979). However, this seems not to be the fact in Campylopus. As shown for the epiphytic species and species of the subgenus Thysanomitrion, these supposed young species have bands of stereids. So the genus Campylopus may be derived from a dicranoid ancestor by evolving hyalocysts (as it might have been the same with the genera of Paraleucobryaceae), but within Campylopus there seems to be support for the opposite evolution, that is reducing the hyalocysts. Transverse sections of the leaf tips of the youngest leaves in Campylopus flexuosus show that the apical cell developes into the median chlorophyllose deuter band. The deuter cells produce abaxial and adaxial hyalocysts (fig. 2a). By this way the transverse section in an early stage of development of a Campylopus leaf resembles that of a mature leaf of Paraleucobryum enerve. Stereids are produced in later stages of development by cell divisions of hyalocysts. In contrast, in young leaves of Dicranum spadiceum Zett., the median deuter cells produce abaxial and adaxial stereids (fig. 2b). The ontogenesis of stereids in Campylopus seems therefore to be different from that in Dicranum. There might also be palaeontological evidence for the fact that hyalocysts in the costae are a plesiomorphous character state. Smoot & Taylor (1986) described a moss from silicified peat of the late Permian of the Antarctic that possessed a multilayered costa. The areolation (figs ) is quite similar to present species of Dicranum. One transverse section (fig. 2) resembles much the structure of a species of Campylopus with a median row of deuter cells, ventral hyalocysts and smaller dorsal, nonstereidal cells. The other transverse section (fig.5), which I would not refer to the same species, shows ventral and dorsal hyalocysts with two median clusters of stereids similar to costa in present species of Brothera or Campylopodiella. Therefore multilayered costae with hyalocysts were already present in the Late Palaeozoic. In conclusion, costae with hyalocysts, erect capsules, a dicranoid peristome with elongate triangular striate teeth split halfway are regarded as plesiomorphic. To the contrary, costae with double stereid bands, asymmetric capsules or filiform peristome teeth are regarded as apomorphic. For the cladistic analysis, the infrageneric categories (subgenus Thysanomitrion, subg. Campylopus with sections Campylopus and Homalocarpus) as proposed by Frahm (1983) have been used along with the subgenus Campylopidulum described later by Vital (1984). The analysis using Henningian cladistics (fig. 3) shows that the subgenera Campylopidulum and Thysanomitrion have the highest number of apomorphous character states, whereas the subgenus
.")
9 99 Campylopus is the most primitive one in which the sect. Homalocarpus turns out to have the most plesiomorphic characters. This does not automatically include any conclusion about the age of these categories. As explained by its rich speciation in SE Asia, the subgenus Thysanomitrion seems to be the youngest branch, indeed. However, the two representatives of subgenus Campylopidulum each with one species in the dry savannah regions of the Neotropics and Africa (Frahm 1986) seem to go back on an origin in the dry climatic periods of the Mesozoic and seem to have developed from the same Gondwanalandic ancestor. At least there was considerable speciation in subg. Campylopus sect. Homalocarpus in the Andes, probably by a spreading of subantarctic ancestors in this mountain chain in Tertiary, although this group is considered here as the most primitive and probably also oldest group. 6. Origin and evolution of the genus It would be worth to speculate about the age of the genus and species and the probable time of its origin. Since there are no species in Campylopus with a pangaean distribution, the genus itself should be younger than 200 Mio years. An indication of the presumed origin of the genus may be a region with the most primitive forms, with a maximum of continuity of the climate through the past geological periods and a continuity of habitats. The most primitive forms according to the cladistic analysis are species of the section Homalocarpus with a center in the subantarctic, where 12 of 14 species belong to this group. A reconstruction of the palaeoclimate allows conclusions whether the climate at any time was humid, either cool or tropical, or arid, and thus less favourable for bryophytes in general or only for adapted species. Furthermore it is important to know to which extent humid climates were present and whether they were continous (allowing migration) or separated by arid regions. An interpretation of geological sediments allows such conclusions in the way that coal deposits indicate humid forests, coral reefs tropical climates, and sandstones, evaporites and Red Beds desert climates. Maps of the coal deposits from different geological periods by Seyfert & Sirkin (1973) show that there must have been a continuous humid climate in the present subantarctic since the late Palaeozoic, which may indicate (together with the high percentage of primitive taxa) an origin of Campylopus along the south coast of Gondwanaland. The next diversification is indicated by dry adapted species with a present wide (gondwanalandic) range from South and Central America to Africa and India. This type of range may go back to the Jurassic period about 150 Mio years ago, when the continents where still connected. Examples are Campylopus pilifer Brid., C. savannarum (C. Müll.) Mitt., or ancestor of species pairs occurring in savannahs such as C. carolinae Grout in Brazil and C. perpusillus Mitt. in Africa. It could be that also mesic forest species such as C. fragilis (Brid.) B.S.G. or C. flexuosus (Hedw.) Brid. were derived in this period by invading the drier interior of Gondwanaland. A reason for this migration and adaptation to drier habitats could be the drainage of swamp forest pressing the species into new habitats (probably combined with a considerable number of extinctions). It could be argued that these widespread ranges are based on relatively recent long distance dispersal. However, this would not explain why Campylopus fragilis, for instance, does not occur in the mountains of SE-Brazil (which should be easily accessible for long distance dispersal) but is replaced there by a local, morphologically different subspecies. It would also not explain why Campylopus flexuosus is not present in SE-Brazil, allthough it has a wide, nearly pantropical range. Therefore these wide ranges seem
 seem to go back on an origin in the dry")
10 100 not to be the result of long distance dispersal but of previously continuous ranges at a time when the continents where still partially connnected. There are some examples for disjunctions of species between SE-Brazil and E- Africa, where they occur in vicariant subspecies or species. Examples are Campylopus trachyblepharon (C. Müll.) Mitt. ssp. trachyblepharon in SE-Brazil and ssp. comatus (Ren. & Card.) J.-P. Frahm in E-Africa, C. julaceus Jaeg. ssp. julaceus and ssp. arbogastii (Ren. & Card.) J.-P. Frahm, C. controversus (Hampe) Jaeg. and C. stenopelma (C. Müll.) Rehm. ex Par. These species must have been present when the South Atlantic had just opened and there was a continuous shoreline stretching from SE-Brazil to Malagasy, about 135 Mio years ago. Separation of the continents lead to isolation and to geographical races. Relicts of the former Campylopus flora of the coastal areas of South America just after the split of Gondwanaland are preserved in SE-Brazil. This region is characterized by 13 endemic species, of which 8 can easily be derived from species with present subantarctic ranges (Frahm in prep.). SE-Brazil had also a continuous humid climate through the past geological periods (Nacib Ab Saber 1982) and thus species could survive here (at least in part) since the Mesozoic. Old species with an age of 100 Mio years or less may be those which are confined to the Neotropics or tropical Africa, where they have a wide range. Examples are Campylopus subcuspidatus (Hampe) Jaeg., C. richardii Brid. and C. lamellinervis (C. Müll.) Mitt. in the Neotropics. Younger species (with an age of 70 Mio years or less) are presumambly those of young Mesozoic islands such as the Caribbean Islands or of mountain massifs of Teriary age. 50% of all neotropical species belong to the andine element and therefore the richest speciation took place during Tertiary. The most recent speciation probably took place at the subspecies level on isolated mountain massifs such as the Serra do Itatiaia in SE-Brazil (Campylopus fragilis (Brid.) B.S.G. ssp. fragiliformis (J.-P. Frahm) J.-P. Frahm) or young volcanic islands such as the Galapagos Islands (C. pilifer Brid. ssp. galapagensis (J.-P. Frahm & Sipman) J.-P. Frahm). There is no convincing indication for speciation during the Quaternary, in spite of strong climatic changes and fluctuation of vegetation belts. It could be that a local endemic like Campylopus cleefii from the colombian páramo and apparently derived from the montane C. arctocarpus (Hornsch.) Mitt. has evolved only in one paramo system, which extended to lower elevations in cold phases of the Pleistocene. However, these fluctuations have probbaly only had an influence on the expansion of present ranges. Based on this distribution-based estimation of the age of the 65 neotropical species of Campylopus, 60% have been evolved in the Tertiary, 30% in Cretaceous, 2% in Jurassic and 8% in the Triassic, demonstrating that the Tertiary speciation was the most important (Frahm, in prep). This should not, however, lead to the conclusion that the number of species of Campylopus has raised through the past 180 Mio years. There were almost certainly numerous extinctions, for example in the Quaternary by lowering or raising of sealevels, lowering and raising of temperatures causing splitting of continuous vegetation belts (like rainforests), and expansion of alpine belts. There are indications of species showing scattered distributions, which probably are extinct in parts of their ranges, like C. subcuspidatus in South America. Other species are confined to extremely small ranges like C. uleanus (C. Müll.) Broth., C. viridatus (C. Müll.) Broth. and C. widgrenii (C. Müll.) Mitt. in SE-Brazil, each known from only 2-4 localities, or have a wide range but with only a few records of small, sterile populations, like
 Rehm. ex Par.")
11 101 C. oerstedianus (C. Müll.) Mitt. a relic tethyan range from central America to Europe, or the Mexican population of C. japonicus Broth. trapped in Mexico as a relict of a previous circum-pacific range. These are the really endangered species and may be the next candidates for extinction. 6. Conclusion Coming back to the statement by Schuster (1984), who called Campylopus a modern and successful genus. Apparently it is successful. This is expressed by the high number of species, and the broad geographical and ecological amplitude. This success is due to anatomical and morphological features, of which the structural plasticity of the costa probably plays the most important role. The genus itself, however, cannot be called a modern genus. The interpretation of the worldwide distribution shows that it probably evolved on the Gondwana continent and therefore has probably been already present in Mesozoic ages. The main speciation, however, may be of Tertiary origin, caused by the uplift of mountain systems and continental drift, and thus diversification is relatively modern. Insofar Campylopus might be called one of the most successful genera of mosses with relatively recent diversification. Parts of the manuscript have been prepared during a stay at the University of Alberta. I wish to thank Dr. D.H.Vitt for providing me with working facilities for this time, support with literature, valuable suggestions and discussions, and also for correcting the English text. Microtome transverse sections were made by M. Campenot. Literature cited: Corley, M.F.V. & J.-P. Frahm Taxonomy and world distribution of Campylopus pyriformis (Schultz) Brid. Journal of Bryology 12: Frahm, J.-P Synopsis of the genus Campylopus in North America North of Mexico. The Bryologist 83: A new infrageneric classification in the genus Campylopus Brid. Journal of the Hattori Botanical Laboratory 54: Phytogeography of European Campylopus species. Proceeding of the Third Meeting of Bryologists from Central and Eastern Europe, Praha, pp The ecological significance of the costal anatomy in the genus Campylopus. Abstracta Botanica 9, Suppl. 2: a. The worldwide range of Campylopus shawii Wils. Journal of Bryology 13: Campylopus Brid. 2. Subg. Campylopidulum Vital. Nova Hedwigia 43: Which factors control the growth of epiphytic bryophytes in tropical rainforests? Symposia Biologica Hungarica 35: a. Oekologische Studien ueber die epiphytische Moosvegetation in Regenwaeldern NO-Perus. Beih. Nova Hedwigia 88: The subantarctic and southern hemispheric species of Campylopus (Dicranaceae), with contributions to the origin and speciation of the genus. Journal Hattori Botanical Laboratory 64: in prep. Origin and distribution of Neotropical species of Campylopus. Frahm, J.-P., M. Giese, M. Padberg, T. Koponen and D.H.Norris Bryophyte flora of the Huon Peninsula, Papua New Guinea. IX. Atractylocarpus, Bryohumbertia, Campylopodium and Campylopus (Dicranaceae, Musci). Acta Botanica Fennica 131: Frey, W. & J.-P. Frahm The twist mechanism in the cygneous setae of the genus Campylopus. Morphology, structure and function. Nova Hedwigia 44: Nacib Ab Saber, A The Paleoclimate and Paleoecology of Brazilian Amazonia. In: G.T.Prance (ed.) Biological Diversification in the Tropics. Proceedings of the Fifth International Symposium of the Association for Tropical Biology held at Macuto Beach, Caracas, Venezuela, 1979, pp Peterson, W A revision of the genera Dicranum and Orthodicranum (Musci) in North America north of Mexico. Ph.D. Thesis Univ. of Alberta. Raven, P.H. & D.I. Axelrod Angiosperm Biogeography and Past Continental Movements. Annals of the Missouri Botanical Garden 61: Robinson, H The structure and significance of the leucobryaceous leaf. Monographs in Systematic Botany from the Missouri Botanical Garden 11: Schuster, R.M Continental Movements, Wallace s Line and Indomalayan-Australasian dispersal of Landplants: Some eclectic concepts. Bot. Rev. 38: 3-86.

Chapter 3 Communities, Biomes, and Ecosystems
 Communities, Biomes, and Ecosystems Section 1: Community Ecology Section 2: Terrestrial Biomes Section 3: Aquatic Ecosystems Click on a lesson name to select. 3.1 Community Ecology Communities A biological
Communities, Biomes, and Ecosystems Section 1: Community Ecology Section 2: Terrestrial Biomes Section 3: Aquatic Ecosystems Click on a lesson name to select. 3.1 Community Ecology Communities A biological
Plate Tectonics Short Study Guide
 Name: Class: Date: Plate Tectonics Short Study Guide Multiple Choice Identify the letter of the choice that best completes the statement or answers the question. 1. The existence of coal beds in Antarctica
Name: Class: Date: Plate Tectonics Short Study Guide Multiple Choice Identify the letter of the choice that best completes the statement or answers the question. 1. The existence of coal beds in Antarctica
Tropical Rainforests in the Pleistocene. Tropical Rainforests in the Pleistocene. Tropical Rainforests in the Pleistocene
 Tropical Rainforests in the Pleistocene tropics stable during Pleistocene? 1 C temperature drop based on 1976 CLIMAP study of warm vs. cold loving forams (vs. 10 C in North Atlantic) Paleothermometers
Tropical Rainforests in the Pleistocene tropics stable during Pleistocene? 1 C temperature drop based on 1976 CLIMAP study of warm vs. cold loving forams (vs. 10 C in North Atlantic) Paleothermometers
REVIEW UNIT 10: ECOLOGY SAMPLE QUESTIONS
 Period Date REVIEW UNIT 10: ECOLOGY SAMPLE QUESTIONS A. Sample Multiple Choice Questions Complete the multiple choice questions to review this unit. 1. All of the following are density-dependent factors
Period Date REVIEW UNIT 10: ECOLOGY SAMPLE QUESTIONS A. Sample Multiple Choice Questions Complete the multiple choice questions to review this unit. 1. All of the following are density-dependent factors
ES Chapter 10 Review. Name: Class: Date: Multiple Choice Identify the choice that best completes the statement or answers the question.
 Name: Class: Date: ES Chapter 10 Review Multiple Choice Identify the choice that best completes the statement or answers the question. 1. Scientists used the pattern of alternating normal and reversed
Name: Class: Date: ES Chapter 10 Review Multiple Choice Identify the choice that best completes the statement or answers the question. 1. Scientists used the pattern of alternating normal and reversed
Communities, Biomes, and Ecosystems
 Communities, Biomes, and Ecosystems Before You Read Before you read the chapter, respond to these statements. 1. Write an A if you agree with the statement. 2. Write a D if you disagree with the statement.
Communities, Biomes, and Ecosystems Before You Read Before you read the chapter, respond to these statements. 1. Write an A if you agree with the statement. 2. Write a D if you disagree with the statement.
Evolution (18%) 11 Items Sample Test Prep Questions
 11 Items Sample Test Prep Questions") Evolution (18%) 11 Items Sample Test Prep Questions Grade 7 (Evolution) 3.a Students know both genetic variation and environmental factors are causes of evolution and diversity of organisms. (pg. 109 Science
Evolution (18%) 11 Items Sample Test Prep Questions Grade 7 (Evolution) 3.a Students know both genetic variation and environmental factors are causes of evolution and diversity of organisms. (pg. 109 Science
THE ECOSYSTEM - Biomes
 Biomes The Ecosystem - Biomes Side 2 THE ECOSYSTEM - Biomes By the end of this topic you should be able to:- SYLLABUS STATEMENT ASSESSMENT STATEMENT CHECK NOTES 2.4 BIOMES 2.4.1 Define the term biome.
Biomes The Ecosystem - Biomes Side 2 THE ECOSYSTEM - Biomes By the end of this topic you should be able to:- SYLLABUS STATEMENT ASSESSMENT STATEMENT CHECK NOTES 2.4 BIOMES 2.4.1 Define the term biome.
Geography affects climate.
 KEY CONCEPT Climate is a long-term weather pattern. BEFORE, you learned The Sun s energy heats Earth s surface unevenly The atmosphere s temperature changes with altitude Oceans affect wind flow NOW, you
KEY CONCEPT Climate is a long-term weather pattern. BEFORE, you learned The Sun s energy heats Earth s surface unevenly The atmosphere s temperature changes with altitude Oceans affect wind flow NOW, you
Flowers; Seeds enclosed in fruit
 Name Class Date Chapter 22 Plant Diversity Section Review 22-1 Reviewing Key Concepts Short Answer On the lines provided, answer the following questions. 1. Describe the main characteristics of plants.
Name Class Date Chapter 22 Plant Diversity Section Review 22-1 Reviewing Key Concepts Short Answer On the lines provided, answer the following questions. 1. Describe the main characteristics of plants.
defined largely by regional variations in climate
 1 Physical Environment: Climate and Biomes EVPP 110 Lecture Instructor: Dr. Largen Fall 2003 2 Climate and Biomes Ecosystem concept physical and biological components of environment are considered as single,
1 Physical Environment: Climate and Biomes EVPP 110 Lecture Instructor: Dr. Largen Fall 2003 2 Climate and Biomes Ecosystem concept physical and biological components of environment are considered as single,
1. The diagram below shows a cross section of sedimentary rock layers.
 1. The diagram below shows a cross section of sedimentary rock layers. Which statement about the deposition of the sediments best explains why these layers have the curved shape shown? 1) Sediments were
1. The diagram below shows a cross section of sedimentary rock layers. Which statement about the deposition of the sediments best explains why these layers have the curved shape shown? 1) Sediments were
Chapter 9: Earth s Past
 Chapter 9: Earth s Past Vocabulary 1. Geologic column 2. Era 3. Period 4. Epoch 5. Evolution 6. Precambrian time 7. Paleozoic era 8. Shield 9. Stromatolite 10. Invertebrate 11. Trilobite 12. Index fossil
Chapter 9: Earth s Past Vocabulary 1. Geologic column 2. Era 3. Period 4. Epoch 5. Evolution 6. Precambrian time 7. Paleozoic era 8. Shield 9. Stromatolite 10. Invertebrate 11. Trilobite 12. Index fossil
Natural Resources and Landscape Survey
 Landscape Info Property Name Address Information Contact Person Relationship to Landscape Email address Phone / Fax Website Address Landscape Type (private/muni/resort, etc.) Former Land Use (if known)
Landscape Info Property Name Address Information Contact Person Relationship to Landscape Email address Phone / Fax Website Address Landscape Type (private/muni/resort, etc.) Former Land Use (if known)
AP BIOLOGY 2007 SCORING GUIDELINES
 AP BIOLOGY 2007 SCORING GUIDELINES Question 3 Compared with other terrestrial biomes, deserts have extremely low productivity. (a) Discuss how temperature, soil composition, and annual precipitation limit
AP BIOLOGY 2007 SCORING GUIDELINES Question 3 Compared with other terrestrial biomes, deserts have extremely low productivity. (a) Discuss how temperature, soil composition, and annual precipitation limit
Deserts, Wind Erosion and Deposition
 Deserts, Wind Erosion and Deposition By definition, a desert has less than 10 in (25 cm) of precipitation per year. Deserts occur at 30 o and 60 o in regions of descending air. Deserts can be hot or cold.
Deserts, Wind Erosion and Deposition By definition, a desert has less than 10 in (25 cm) of precipitation per year. Deserts occur at 30 o and 60 o in regions of descending air. Deserts can be hot or cold.
Lecture 10 Friday, March 20, 2009
 Lecture 10 Friday, March 20, 2009 Reproductive isolating mechanisms Prezygotic barriers: Anything that prevents mating and fertilization is a prezygotic mechanism. Habitat isolation, behavioral isolation,
Lecture 10 Friday, March 20, 2009 Reproductive isolating mechanisms Prezygotic barriers: Anything that prevents mating and fertilization is a prezygotic mechanism. Habitat isolation, behavioral isolation,
Name: DUE: May 2, 2013 Ms. Galaydick. Geologic Time Scale Era Period End date (in millions of years) Cenozoic Quaternary present
 Cenozoic Quaternary present") Name: DUE: May 2, 2013 Ms. Galaydick Objective: Use the diagrams to answer the questions for each set: USING SCIENCE SKILLS PART #1 Geologic Time Scale Era Period End date (in millions of years) Cenozoic
Name: DUE: May 2, 2013 Ms. Galaydick Objective: Use the diagrams to answer the questions for each set: USING SCIENCE SKILLS PART #1 Geologic Time Scale Era Period End date (in millions of years) Cenozoic
Chapter 18 Introduction to. A f r i c a
 Chapter 18 Introduction to A f r i c a Ch. 18:1 Landforms & Resources 1. Africa s shape & landforms are the result of its location in the southern part of the ancient supercontinent of. Pangaea Over thousands
Chapter 18 Introduction to A f r i c a Ch. 18:1 Landforms & Resources 1. Africa s shape & landforms are the result of its location in the southern part of the ancient supercontinent of. Pangaea Over thousands
Rocks and Plate Tectonics
 Name: Class: _ Date: _ Rocks and Plate Tectonics Multiple Choice Identify the choice that best completes the statement or answers the question. 1. What is a naturally occurring, solid mass of mineral or
Name: Class: _ Date: _ Rocks and Plate Tectonics Multiple Choice Identify the choice that best completes the statement or answers the question. 1. What is a naturally occurring, solid mass of mineral or
OCEANS AND AQUATIC ECOSYSTEMS - Vol. II - Conservation of Island Plant Populations and Communities - Steve Waldren
 CONSERVATION OF ISLAND PLANT POPULATIONS AND COMMUNITIES Steve Waldren Trinity College Botanic Garden, Dublin, Ireland Keywords: conservation; deterministic threats; Easter Island; endangered species;
CONSERVATION OF ISLAND PLANT POPULATIONS AND COMMUNITIES Steve Waldren Trinity College Botanic Garden, Dublin, Ireland Keywords: conservation; deterministic threats; Easter Island; endangered species;
Unit 2 Lesson 4 The Geologic Time Scale. Copyright Houghton Mifflin Harcourt Publishing Company
 Once Upon a Time How have geologists described the rate of geologic change? Geology is the scientific study of the origin, history, and structure of Earth and the processes that shape it. Early geologists
Once Upon a Time How have geologists described the rate of geologic change? Geology is the scientific study of the origin, history, and structure of Earth and the processes that shape it. Early geologists
Geologic Time Scale Notes
 Name: Date: Period: 1 2 3 4 5 6 7 Notes Essential Question: What is the geologic time scale? Vocabulary: Geology: the scientific study of the origin, history, and structure of Earth and the processes that
Name: Date: Period: 1 2 3 4 5 6 7 Notes Essential Question: What is the geologic time scale? Vocabulary: Geology: the scientific study of the origin, history, and structure of Earth and the processes that
California Standards Grades 9 12 Boardworks 2009 Science Contents Standards Mapping
 California Standards Grades 912 Boardworks 2009 Science Contents Standards Mapping Earth Sciences Earth s Place in the Universe 1. Astronomy and planetary exploration reveal the solar system s structure,
California Standards Grades 912 Boardworks 2009 Science Contents Standards Mapping Earth Sciences Earth s Place in the Universe 1. Astronomy and planetary exploration reveal the solar system s structure,
BIOL 1030 TOPIC 5 LECTURE NOTES TOPIC 5: SEEDLESS VASCULAR PLANTS (CH. 29)
") TOPIC 5: SEEDLESS VASCULAR PLANTS (CH. 29) I. Vascular Plants (overview) plants with xylem and phloem 7 or 9 living phyla, depending on who you talk to able to dominate most terrestrial habitats because
TOPIC 5: SEEDLESS VASCULAR PLANTS (CH. 29) I. Vascular Plants (overview) plants with xylem and phloem 7 or 9 living phyla, depending on who you talk to able to dominate most terrestrial habitats because
Geologic History Review
 1. The climate that existed in an area during the early Paleozoic Era can best be determined by studying (1) the present climate of the area (2) recorded climate data of the area since 1700 (3) present
1. The climate that existed in an area during the early Paleozoic Era can best be determined by studying (1) the present climate of the area (2) recorded climate data of the area since 1700 (3) present
Continents join together and split apart.
 KEY CONCEPT Continents change position over time. BEFORE, you learned Earth s main layers are the core, the mantle, and the crust The lithosphere and asthenosphere are the topmost layers of Earth The lithosphere
KEY CONCEPT Continents change position over time. BEFORE, you learned Earth s main layers are the core, the mantle, and the crust The lithosphere and asthenosphere are the topmost layers of Earth The lithosphere
Key Idea 2: Ecosystems
 Key Idea 2: Ecosystems Ecosystems An ecosystem is a living community of plants and animals sharing an environment with non-living elements such as climate and soil. An example of a small scale ecosystem
Key Idea 2: Ecosystems Ecosystems An ecosystem is a living community of plants and animals sharing an environment with non-living elements such as climate and soil. An example of a small scale ecosystem
Exhibit Inquiry. Rainforest. Aug 11
 Exhibit Inquiry Exhibit Inquiry Have students look for the following exhibits related to living things during their visit to the Ontario Science Centre: Where to go: (Level 6) What it's about: Tropical
Exhibit Inquiry Exhibit Inquiry Have students look for the following exhibits related to living things during their visit to the Ontario Science Centre: Where to go: (Level 6) What it's about: Tropical
Review of the Late Pleistocene Soricidae (Mammalia) fauna of the Vaskapu Cave (North Hungary)
 fauna of the Vaskapu Cave (North Hungary)") Hantkeniana 8, 163 169, Budapest, 2013 Review of the Late Pleistocene Soricidae (Mammalia) fauna of the Vaskapu Cave (North Hungary) Lukács MÉSZÁROS 1 (with 2 figures, 2 tables and 1 plate) The summary
Hantkeniana 8, 163 169, Budapest, 2013 Review of the Late Pleistocene Soricidae (Mammalia) fauna of the Vaskapu Cave (North Hungary) Lukács MÉSZÁROS 1 (with 2 figures, 2 tables and 1 plate) The summary
Physiography, Geography and Climate of Latin America (Lecture 3)
") Physiography, Geography and Climate of Latin America (Lecture 3) Natural Landmarks in Latin America World's longest and second highest mountain range, and the world's highest active volcanoes. Biggest
Physiography, Geography and Climate of Latin America (Lecture 3) Natural Landmarks in Latin America World's longest and second highest mountain range, and the world's highest active volcanoes. Biggest
A Few Facts about Antarctica
 A Few Facts about Antarctica Antarctica is the continent that surrounds the South Pole, the southernmost point at the bottom of the earth. Antarctica is a continent because it is land that is covered by
A Few Facts about Antarctica Antarctica is the continent that surrounds the South Pole, the southernmost point at the bottom of the earth. Antarctica is a continent because it is land that is covered by
Teaching the Dynamic Earth Plate Tectonics Interactive Master sheets
 Teaching the Dynamic Earth Plate Tectonics Interactive Master sheets Contents Page Master Sheets for continental jigsaws Gondwanaland continents (blank outlines)...2 Four continents, showing gaps and overlap
Teaching the Dynamic Earth Plate Tectonics Interactive Master sheets Contents Page Master Sheets for continental jigsaws Gondwanaland continents (blank outlines)...2 Four continents, showing gaps and overlap
BALBHARATI PUBLIC SCHOOL, PITAMPURA,NEW DELHI COMPREHENSIVE NOTES CHAPTER-6 HUMAN RESOURCES CLASS-VIII SUBJECT-GEOGRAPHY FINAL TERM
 BALBHARATI PUBLIC SCHOOL, PITAMPURA,NEW DELHI COMPREHENSIVE NOTES CHAPTER-6 HUMAN RESOURCES CLASS-VIII SUBJECT-GEOGRAPHY FINAL TERM Q1. Why are people considered as a resource? A1. People are a nation
BALBHARATI PUBLIC SCHOOL, PITAMPURA,NEW DELHI COMPREHENSIVE NOTES CHAPTER-6 HUMAN RESOURCES CLASS-VIII SUBJECT-GEOGRAPHY FINAL TERM Q1. Why are people considered as a resource? A1. People are a nation
A Morphological Study On Endemic Malabaila lasiocarpa Boiss. (Apiaceae) From Bingol (Turkey)
 From Bingol (Turkey)") Araştırma Makalesi/Research Article A Morphological Study On Endemic Malabaila lasiocarpa Boiss. (Apiaceae) From Bingol (Turkey) Ömer KILIÇ 1 Abstract- In this study morphological characters of Malabaila
Araştırma Makalesi/Research Article A Morphological Study On Endemic Malabaila lasiocarpa Boiss. (Apiaceae) From Bingol (Turkey) Ömer KILIÇ 1 Abstract- In this study morphological characters of Malabaila
What are the controls for calcium carbonate distribution in marine sediments?
 Lecture 14 Marine Sediments (1) The CCD is: (a) the depth at which no carbonate secreting organisms can live (b) the depth at which seawater is supersaturated with respect to calcite (c) the depth at which
Lecture 14 Marine Sediments (1) The CCD is: (a) the depth at which no carbonate secreting organisms can live (b) the depth at which seawater is supersaturated with respect to calcite (c) the depth at which
3 The Mesozoic and Cenozoic Eras
 CHAPTER 9 3 The Mesozoic and Cenozoic Eras SECTION A View of Earth s Past KEY IDEAS As you read this section, keep these questions in mind: What were the periods of the Mesozoic and Cenozoic Eras? What
CHAPTER 9 3 The Mesozoic and Cenozoic Eras SECTION A View of Earth s Past KEY IDEAS As you read this section, keep these questions in mind: What were the periods of the Mesozoic and Cenozoic Eras? What
Tropical Rainforest. Abiotic Factors Amount of Water, Sunlight, Soil, Precipitation
 World Biomes A biome is an area of land that shares similar temperatures and precipitation. The observation of the temperature and precipitation over a period time make up a biome s climate. Each biome
World Biomes A biome is an area of land that shares similar temperatures and precipitation. The observation of the temperature and precipitation over a period time make up a biome s climate. Each biome
SLOW ONSET EVENTS. climate change impacts on BIODIVERSITY
 Regional Gateway for Technology Transfer and Climate Change Action in Latin America and the Caribbean (ROLAC UNEP) Characterizing and addressing SLOW ONSET EVENTS climate change impacts on BIODIVERSITY
Regional Gateway for Technology Transfer and Climate Change Action in Latin America and the Caribbean (ROLAC UNEP) Characterizing and addressing SLOW ONSET EVENTS climate change impacts on BIODIVERSITY
Name Class Date WHAT I KNOW. about how organisms have changed. grown in complexity over time.
 History of Life Evolution Q: How do fossils help biologists understand the history of life on Earth? 19.1 How do scientists use fossils to study Earth s history? WHAT I KNOW SAMPLE ANSWER: Fossils give
History of Life Evolution Q: How do fossils help biologists understand the history of life on Earth? 19.1 How do scientists use fossils to study Earth s history? WHAT I KNOW SAMPLE ANSWER: Fossils give
Data Quality Working Group
 Data Quality Working Group In order to get an overview of the issues that have been discussed so far, I have summarized them a little and added some links and references. The interesting discussion is
Data Quality Working Group In order to get an overview of the issues that have been discussed so far, I have summarized them a little and added some links and references. The interesting discussion is
What Causes Climate? Use Target Reading Skills
 Climate and Climate Change Name Date Class Climate and Climate Change Guided Reading and Study What Causes Climate? This section describes factors that determine climate, or the average weather conditions
Climate and Climate Change Name Date Class Climate and Climate Change Guided Reading and Study What Causes Climate? This section describes factors that determine climate, or the average weather conditions
Geol 101: Physical Geology PAST EXAM QUESTIONS LECTURE 4: PLATE TECTONICS II
 Geol 101: Physical Geology PAST EXAM QUESTIONS LECTURE 4: PLATE TECTONICS II 4. Which of the following statements about paleomagnetism at spreading ridges is FALSE? A. there is a clear pattern of paleomagnetic
Geol 101: Physical Geology PAST EXAM QUESTIONS LECTURE 4: PLATE TECTONICS II 4. Which of the following statements about paleomagnetism at spreading ridges is FALSE? A. there is a clear pattern of paleomagnetic
10B Plant Systems Guided Practice
 10B Plant Systems Guided Practice Reproduction Station 1 1. Observe Plant A. Locate the following parts of the flower: stamen, stigma, style, ovary. 2. Draw and label the parts of a flower (listed above)
10B Plant Systems Guided Practice Reproduction Station 1 1. Observe Plant A. Locate the following parts of the flower: stamen, stigma, style, ovary. 2. Draw and label the parts of a flower (listed above)
Regents Questions: Plate Tectonics
 Earth Science Regents Questions: Plate Tectonics Name: Date: Period: August 2013 Due Date: 17 Compared to the oceanic crust, the continental crust is (1) less dense and more basaltic (3) more dense and
Earth Science Regents Questions: Plate Tectonics Name: Date: Period: August 2013 Due Date: 17 Compared to the oceanic crust, the continental crust is (1) less dense and more basaltic (3) more dense and
Practice Questions 1: Evolution
 Practice Questions 1: Evolution 1. Which concept is best illustrated in the flowchart below? A. natural selection B. genetic manipulation C. dynamic equilibrium D. material cycles 2. The diagram below
Practice Questions 1: Evolution 1. Which concept is best illustrated in the flowchart below? A. natural selection B. genetic manipulation C. dynamic equilibrium D. material cycles 2. The diagram below
Week 1. Week 2. Week 3
 Week 1 1. What US city has the largest population? 2. Where is Aachen? 3. What is the capitol of Florida? 4. What is the longest mountain range in Spain? 5. What countries border Equador? Week 2 1. What
Week 1 1. What US city has the largest population? 2. Where is Aachen? 3. What is the capitol of Florida? 4. What is the longest mountain range in Spain? 5. What countries border Equador? Week 2 1. What
11A Plate Tectonics. What is plate tectonics? Setting up. Materials
 11A Plate Tectonics What is plate tectonics? Earth s crust plus the upper mantle forms the lithosphere. Earth s lithosphere is broken in a number of different pieces. How these pieces move and interact
11A Plate Tectonics What is plate tectonics? Earth s crust plus the upper mantle forms the lithosphere. Earth s lithosphere is broken in a number of different pieces. How these pieces move and interact
Name Period 4 th Six Weeks Notes 2015 Weather
 Name Period 4 th Six Weeks Notes 2015 Weather Radiation Convection Currents Winds Jet Streams Energy from the Sun reaches Earth as electromagnetic waves This energy fuels all life on Earth including the
Name Period 4 th Six Weeks Notes 2015 Weather Radiation Convection Currents Winds Jet Streams Energy from the Sun reaches Earth as electromagnetic waves This energy fuels all life on Earth including the
Biology 1406 - Notes for exam 5 - Population genetics Ch 13, 14, 15
 Biology 1406 - Notes for exam 5 - Population genetics Ch 13, 14, 15 Species - group of individuals that are capable of interbreeding and producing fertile offspring; genetically similar 13.7, 14.2 Population
Biology 1406 - Notes for exam 5 - Population genetics Ch 13, 14, 15 Species - group of individuals that are capable of interbreeding and producing fertile offspring; genetically similar 13.7, 14.2 Population
Orbital-Scale Climate Change
 Orbital-Scale Climate Change Climate Needed for Ice Age Warm winter and non-frozen oceans so lots of evaporation and snowfall Cool summer so that ice does not melt Ice Age Model When ice growing ocean
Orbital-Scale Climate Change Climate Needed for Ice Age Warm winter and non-frozen oceans so lots of evaporation and snowfall Cool summer so that ice does not melt Ice Age Model When ice growing ocean
SOUTH AMERICA CONTENTS. What s in This Book... 2. Section 1: South America in the World... 3. Section 2: Political Divisions of South America...
 SOUTH CONTENTS What s in This Book 2 Section 1: 3 Section 2: Political Divisions of 1 Section 3: Physical Features of 41 Section 4: Valuable Resources of 67 Section : n Culture 89 Section 6: Assessment
SOUTH CONTENTS What s in This Book 2 Section 1: 3 Section 2: Political Divisions of 1 Section 3: Physical Features of 41 Section 4: Valuable Resources of 67 Section : n Culture 89 Section 6: Assessment
6. Base your answer to the following question on the graph below, which shows the average monthly temperature of two cities A and B.
 1. Which single factor generally has the greatest effect on the climate of an area on the Earth's surface? 1) the distance from the Equator 2) the extent of vegetative cover 3) the degrees of longitude
1. Which single factor generally has the greatest effect on the climate of an area on the Earth's surface? 1) the distance from the Equator 2) the extent of vegetative cover 3) the degrees of longitude
Earth Sciences -- Grades 9, 10, 11, and 12. California State Science Content Standards. Mobile Climate Science Labs
 Earth Sciences -- Grades 9, 10, 11, and 12 California State Science Content Standards Covered in: Hands-on science labs, demonstrations, & activities. Investigation and Experimentation. Lesson Plans. Presented
Earth Sciences -- Grades 9, 10, 11, and 12 California State Science Content Standards Covered in: Hands-on science labs, demonstrations, & activities. Investigation and Experimentation. Lesson Plans. Presented
Alfred Wegener s Theory of Continental Drift Became Modern Plate Tectonics. Wegener in Greenland about 1912. He froze to death there in 1930.
 Alfred Wegener s Theory of Continental Drift Became Modern Plate Tectonics Wegener in Greenland about 1912. He froze to death there in 1930. Science is self correcting. The Scientific Method The history
Alfred Wegener s Theory of Continental Drift Became Modern Plate Tectonics Wegener in Greenland about 1912. He froze to death there in 1930. Science is self correcting. The Scientific Method The history
WEATHER, CLIMATE AND ADAPTATIONS OF ANIMALS TO CLIMATE
 7 WEATHER, CLIMATE AND ADAPTATIONS OF ANIMALS TO CLIMATE TEXTBOOK QUESTIONS AND ANSWERS Q.1. Why weather changes so frequently? Ans. All changes in the weather are caused by the sun. The movement of the
7 WEATHER, CLIMATE AND ADAPTATIONS OF ANIMALS TO CLIMATE TEXTBOOK QUESTIONS AND ANSWERS Q.1. Why weather changes so frequently? Ans. All changes in the weather are caused by the sun. The movement of the
Stage 4. Geography. Blackline Masters. By Karen Devine
 1 Devine Educational Consultancy Services Stage 4 Geography Blackline Masters By Karen Devine Updated January 2010 2 This book is intended for the exclusive use in NSW Secondary Schools. It is meant to
1 Devine Educational Consultancy Services Stage 4 Geography Blackline Masters By Karen Devine Updated January 2010 2 This book is intended for the exclusive use in NSW Secondary Schools. It is meant to
The Earth System. The geosphere is the solid Earth that includes the continental and oceanic crust as well as the various layers of Earth s interior.
 The Earth System The atmosphere is the gaseous envelope that surrounds Earth. It consists of a mixture of gases composed primarily of nitrogen, oxygen, carbon dioxide, and water vapor. The atmosphere and
The Earth System The atmosphere is the gaseous envelope that surrounds Earth. It consists of a mixture of gases composed primarily of nitrogen, oxygen, carbon dioxide, and water vapor. The atmosphere and
Biomes An Overview of Ecology Biomes Freshwater Biomes
 Biomes An Overview of Ecology Ecology is the scientific study of the interactions between organisms and their environments. Ecology can be divided into four increasingly comprehensive levels: Organismal
Biomes An Overview of Ecology Ecology is the scientific study of the interactions between organisms and their environments. Ecology can be divided into four increasingly comprehensive levels: Organismal
Scope and Sequence Interactive Science grades 6-8
 Science and Technology Chapter 1. What Is Science? 1. Science and the Natural World 2.Thinking Like a Scientist 3. Scientific Inquiry Scope and Sequence Interactive Science grades 6-8 Chapter 2. Science,
Science and Technology Chapter 1. What Is Science? 1. Science and the Natural World 2.Thinking Like a Scientist 3. Scientific Inquiry Scope and Sequence Interactive Science grades 6-8 Chapter 2. Science,
WEATHERING, EROSION, AND DEPOSITION PRACTICE TEST. Which graph best shows the relative stream velocities across the stream from A to B?
 NAME DATE WEATHERING, EROSION, AND DEPOSITION PRACTICE TEST 1. The diagram below shows a meandering stream. Measurements of stream velocity were taken along straight line AB. Which graph best shows the
NAME DATE WEATHERING, EROSION, AND DEPOSITION PRACTICE TEST 1. The diagram below shows a meandering stream. Measurements of stream velocity were taken along straight line AB. Which graph best shows the
The concepts developed in this standard include the following: Oceans cover about 70% of the surface of the Earth.
 Name Date Grade 5 SOL 5.6 Review Oceans Made by SOLpass - www.solpass.org [email protected] Reproduction is permitted for SOLpass subscribers only. The concepts developed in this standard include
Name Date Grade 5 SOL 5.6 Review Oceans Made by SOLpass - www.solpass.org [email protected] Reproduction is permitted for SOLpass subscribers only. The concepts developed in this standard include
Plate Tectonics. Introduction. Boundaries between crustal plates
 Plate Tectonics KEY WORDS: continental drift, seafloor spreading, plate tectonics, mid ocean ridge (MOR) system, spreading center, rise, divergent plate boundary, subduction zone, convergent plate boundary,
Plate Tectonics KEY WORDS: continental drift, seafloor spreading, plate tectonics, mid ocean ridge (MOR) system, spreading center, rise, divergent plate boundary, subduction zone, convergent plate boundary,
 Stanley Price, M. 1969. The bongo of the Cherengani Hills. 10: 109-111. Stanley Price, M. 1978. Fringe-eared oryx on a Kenya ranch. 14: 370-373. Stelfox, B. 1984. Wildlife ranching. 15-l 9. Stelfox, J.G.;
Stanley Price, M. 1969. The bongo of the Cherengani Hills. 10: 109-111. Stanley Price, M. 1978. Fringe-eared oryx on a Kenya ranch. 14: 370-373. Stelfox, B. 1984. Wildlife ranching. 15-l 9. Stelfox, J.G.;
Plant Structure, Growth, and Development. Chapter 35
 Plant Structure, Growth, and Development Chapter 35 PLANTS developmental plasticity = ability of plant to alter form to respond to environment Biological heirarchy Cell basic unit of life Tissue group
Plant Structure, Growth, and Development Chapter 35 PLANTS developmental plasticity = ability of plant to alter form to respond to environment Biological heirarchy Cell basic unit of life Tissue group
The Ice Age By: Sue Peterson
 www.k5learning.com Objective sight words (pulses, intermittent, isotopes, chronicle, methane, tectonic plates, volcanism, configurations, land-locked, erratic); concepts (geological evidence and specific
www.k5learning.com Objective sight words (pulses, intermittent, isotopes, chronicle, methane, tectonic plates, volcanism, configurations, land-locked, erratic); concepts (geological evidence and specific
How Did These Ocean Features and Continental Margins Form?
 298 10.14 INVESTIGATION How Did These Ocean Features and Continental Margins Form? The terrain below contains various features on the seafloor, as well as parts of three continents. Some general observations
298 10.14 INVESTIGATION How Did These Ocean Features and Continental Margins Form? The terrain below contains various features on the seafloor, as well as parts of three continents. Some general observations
Chapter 3: Climate and Climate Change Answers
 Chapter 3: Climate and Climate Change Answers Section A: Climate 1. (a) Explain what each of the following means: (4 x 1 mark) (i) climate the average weather of an area over a 25 30 year period (ii) maritime
Chapter 3: Climate and Climate Change Answers Section A: Climate 1. (a) Explain what each of the following means: (4 x 1 mark) (i) climate the average weather of an area over a 25 30 year period (ii) maritime
Communities, Biomes, and Ecosystems
 Chapter Test A CHAPTER 3 Communities, Biomes, and Ecosystems Part A: Multiple Choice In the space at the left, write the letter of the term or phrase that best answers each question Part B: Matching 1
Chapter Test A CHAPTER 3 Communities, Biomes, and Ecosystems Part A: Multiple Choice In the space at the left, write the letter of the term or phrase that best answers each question Part B: Matching 1
Functional Biology of Plants
 Brochure More information from http://www.researchandmarkets.com/reports/2252012/ Functional Biology of Plants Description: Functional Biology of Plants provides students and researchers with a clearly
Brochure More information from http://www.researchandmarkets.com/reports/2252012/ Functional Biology of Plants Description: Functional Biology of Plants provides students and researchers with a clearly
Sullivan s Island Bird Banding and Environmental Education Program. Sarah Harper Díaz, MA and Jennifer Tyrrell, MS
 Sullivan s Island Bird Banding and Environmental Education Program Sarah Harper Díaz, MA and Jennifer Tyrrell, MS I. Introduction The Sullivan s Island Bird Banding and Environmental Education Program
Sullivan s Island Bird Banding and Environmental Education Program Sarah Harper Díaz, MA and Jennifer Tyrrell, MS I. Introduction The Sullivan s Island Bird Banding and Environmental Education Program
Introduction to Plants
 Introduction to Plants Unity and Diversity of Life Q: What are the five main groups of plants, and how have four of these groups adapted to life on land? 22.1 What are of plants? WHAT I KNOW SAMPLE ANSWER:
Introduction to Plants Unity and Diversity of Life Q: What are the five main groups of plants, and how have four of these groups adapted to life on land? 22.1 What are of plants? WHAT I KNOW SAMPLE ANSWER:
The Fossil Record and Geologic Time Scale
 Two Conceptions of Earth History: Catastrophism Assumption: Great Effects Require Great Causes Earth History Dominated by Violent Events Uniformitarianism Assumption: We Can Use Cause And Effect to Determine
Two Conceptions of Earth History: Catastrophism Assumption: Great Effects Require Great Causes Earth History Dominated by Violent Events Uniformitarianism Assumption: We Can Use Cause And Effect to Determine
Terms of Reference For First Joint Mission for the Pilot Programme on Climate Resilience (PPCR) In St. Vincent and the Grenadines
 In St. Vincent and the Grenadines") Terms of Reference For First Joint Mission for the Pilot Programme on Climate Resilience (PPCR) In St. Vincent and the Grenadines 1 Glossary of Terms and Abbreviations CIF IDB MTESP NEMO NESDC NESDP NGO
Terms of Reference For First Joint Mission for the Pilot Programme on Climate Resilience (PPCR) In St. Vincent and the Grenadines 1 Glossary of Terms and Abbreviations CIF IDB MTESP NEMO NESDC NESDP NGO
Chapter Overview. Seasons. Earth s Seasons. Distribution of Solar Energy. Solar Energy on Earth. CHAPTER 6 Air-Sea Interaction
 Chapter Overview CHAPTER 6 Air-Sea Interaction The atmosphere and the ocean are one independent system. Earth has seasons because of the tilt on its axis. There are three major wind belts in each hemisphere.
Chapter Overview CHAPTER 6 Air-Sea Interaction The atmosphere and the ocean are one independent system. Earth has seasons because of the tilt on its axis. There are three major wind belts in each hemisphere.
Complete tests for CO 2 and H 2 Link observations of acid reactions to species
 Acids and Bases 1. Name common acids and bases found at home and at school 2. Use formulae for common acids and bases 3. Give examples of the uses of acids and bases 4. State that all solutions are acidic,
Acids and Bases 1. Name common acids and bases found at home and at school 2. Use formulae for common acids and bases 3. Give examples of the uses of acids and bases 4. State that all solutions are acidic,
Recent ostracods from the Azores archipelago
 Joannea Geol. Paläont. 11: 132-136 (2011) Recent ostracods from the Azores archipelago Ricardo P. MEIRELES, Antonio FRIAS MARTINS & Sérgio ÁVILA The Azores is an archipelago in the Atlantic Ocean between
Joannea Geol. Paläont. 11: 132-136 (2011) Recent ostracods from the Azores archipelago Ricardo P. MEIRELES, Antonio FRIAS MARTINS & Sérgio ÁVILA The Azores is an archipelago in the Atlantic Ocean between
Plants, like all other living organisms have basic needs: a source of nutrition (food),
,") LEARNING FROM LEAVES: A LOOK AT LEAF SIZE Grades 3 6 I. Introduction Plants, like all other living organisms have basic needs: a source of nutrition (food), water, space in which to live, air, and optimal
LEARNING FROM LEAVES: A LOOK AT LEAF SIZE Grades 3 6 I. Introduction Plants, like all other living organisms have basic needs: a source of nutrition (food), water, space in which to live, air, and optimal
XVI. Science and Technology/Engineering, Grade 5
 XVI. Science and Technology/Engineering, Grade 5 Grade 5 Science and Technology/Engineering Test The spring 2012 grade 5 Science and Technology/Engineering test was based on learning standards in the four
XVI. Science and Technology/Engineering, Grade 5 Grade 5 Science and Technology/Engineering Test The spring 2012 grade 5 Science and Technology/Engineering test was based on learning standards in the four
Summary. 16 1 Genes and Variation. 16 2 Evolution as Genetic Change. Name Class Date
 Chapter 16 Summary Evolution of Populations 16 1 Genes and Variation Darwin s original ideas can now be understood in genetic terms. Beginning with variation, we now know that traits are controlled by
Chapter 16 Summary Evolution of Populations 16 1 Genes and Variation Darwin s original ideas can now be understood in genetic terms. Beginning with variation, we now know that traits are controlled by
What is the human impact on biodiversity? -----------------------
 Biology and Society Unit Seven: The Human Impact Topic Three: Biodiversity What is the human impact on biodiversity? How many species are there? We currently have identified and named approximately 1.5
Biology and Society Unit Seven: The Human Impact Topic Three: Biodiversity What is the human impact on biodiversity? How many species are there? We currently have identified and named approximately 1.5
TECTONICS ASSESSMENT
 Tectonics Assessment / 1 TECTONICS ASSESSMENT 1. Movement along plate boundaries produces A. tides. B. fronts. C. hurricanes. D. earthquakes. 2. Which of the following is TRUE about the movement of continents?
Tectonics Assessment / 1 TECTONICS ASSESSMENT 1. Movement along plate boundaries produces A. tides. B. fronts. C. hurricanes. D. earthquakes. 2. Which of the following is TRUE about the movement of continents?
Advice For the multiple-choice questions, completely fill in the circle alongside the appropriate answer(s).
.") SPECIMEN ASSESSMENT MATERIAL GCSE GEOGRAPHY Paper 1 Living with the physical environment Specimen Materials For this paper you must have: a pencil a ruler. Time allowed: 1 hour 30 minutes Instructions
SPECIMEN ASSESSMENT MATERIAL GCSE GEOGRAPHY Paper 1 Living with the physical environment Specimen Materials For this paper you must have: a pencil a ruler. Time allowed: 1 hour 30 minutes Instructions
Amherst County Public Schools. AP Environmental Science Curriculum Pacing Guide. College Board AP Environmental Science Site
 Amherst County Public Schools AP Environmental Science Curriculum Pacing Guide College Board AP Environmental Science Site REV: 8/12 1 st 9 weeks AP Objectives Energy Resources and Consumption A. Energy
Amherst County Public Schools AP Environmental Science Curriculum Pacing Guide College Board AP Environmental Science Site REV: 8/12 1 st 9 weeks AP Objectives Energy Resources and Consumption A. Energy
TIDES. 1. Tides are the regular rise and fall of sea level that occurs either once a day (every 24.8 hours) or twice a day (every 12.4 hours).
 or twice a day (every 12.4 hours).") TIDES What causes tides? How are tides predicted? 1. Tides are the regular rise and fall of sea level that occurs either once a day (every 24.8 hours) or twice a day (every 12.4 hours). Tides are waves
TIDES What causes tides? How are tides predicted? 1. Tides are the regular rise and fall of sea level that occurs either once a day (every 24.8 hours) or twice a day (every 12.4 hours). Tides are waves
Evidence for evolution factsheet
 The theory of evolution by natural selection is supported by a great deal of evidence. Fossils Fossils are formed when organisms become buried in sediments, causing little decomposition of the organism.
The theory of evolution by natural selection is supported by a great deal of evidence. Fossils Fossils are formed when organisms become buried in sediments, causing little decomposition of the organism.
SECOND GRADE PLATE TECTONICS 1 WEEK LESSON PLANS AND ACTIVITIES
 SECOND GRADE PLATE TECTONICS 1 WEEK LESSON PLANS AND ACTIVITIES PLATE TECTONIC CYCLE OVERVIEW OF SECOND GRADE VOLCANOES WEEK 1. PRE: Investigating the parts of a volcano. LAB: Comparing the parts of a
SECOND GRADE PLATE TECTONICS 1 WEEK LESSON PLANS AND ACTIVITIES PLATE TECTONIC CYCLE OVERVIEW OF SECOND GRADE VOLCANOES WEEK 1. PRE: Investigating the parts of a volcano. LAB: Comparing the parts of a
Hurricanes. Characteristics of a Hurricane
 Hurricanes Readings: A&B Ch. 12 Topics 1. Characteristics 2. Location 3. Structure 4. Development a. Tropical Disturbance b. Tropical Depression c. Tropical Storm d. Hurricane e. Influences f. Path g.
Hurricanes Readings: A&B Ch. 12 Topics 1. Characteristics 2. Location 3. Structure 4. Development a. Tropical Disturbance b. Tropical Depression c. Tropical Storm d. Hurricane e. Influences f. Path g.
Productivity in Cloud Forests in the Andes
 Productivity in Cloud Forests in the Andes Carbon cycle measurements and model results from JULES Toby Marthews http://marthews.tripod.com Leeds, 10th June 2010 at the Oxford University Centre for the
Productivity in Cloud Forests in the Andes Carbon cycle measurements and model results from JULES Toby Marthews http://marthews.tripod.com Leeds, 10th June 2010 at the Oxford University Centre for the
In this lesson, students will identify a local plant community and make a variety of
 MAKING COMMUNITY MEASUREMENTS: ABIOTIC FACTORS Grades 3 6 I. Introduction In this lesson, students will identify a local plant community and make a variety of measurements, preferably during two different
MAKING COMMUNITY MEASUREMENTS: ABIOTIC FACTORS Grades 3 6 I. Introduction In this lesson, students will identify a local plant community and make a variety of measurements, preferably during two different
Plate Tectonics: Big Ideas. Plate Tectonics. Plate Tectonics. The unifying concept of the Earth sciences.
 Plate Tectonics: Big Ideas Our understanding of Earth is continuously refined. Earth s systems are dynamic; they continually react to changing influences from geological, hydrological, physical, chemical,
Plate Tectonics: Big Ideas Our understanding of Earth is continuously refined. Earth s systems are dynamic; they continually react to changing influences from geological, hydrological, physical, chemical,
PLANT EVOLUTION DISPLAY Handout
 PLANT EVOLUTION DISPLAY Handout Name: TA and Section time Welcome to UCSC Greenhouses. This sheet explains a few botanical facts about plant reproduction that will help you through the display and handout.
PLANT EVOLUTION DISPLAY Handout Name: TA and Section time Welcome to UCSC Greenhouses. This sheet explains a few botanical facts about plant reproduction that will help you through the display and handout.
Tectonic plates have different boundaries.
 KEY CONCEPT Plates move apart. BEFORE, you learned The continents join and break apart The sea floor provides evidence that tectonic plates move The theory of plate tectonics helps explain how the plates
KEY CONCEPT Plates move apart. BEFORE, you learned The continents join and break apart The sea floor provides evidence that tectonic plates move The theory of plate tectonics helps explain how the plates
6.E.2.2 Plate Tectonics, Earthquakes and Volcanoes
 Name: Date: 1. The road shown below was suddenly broken by a natural event. 3. The convergence of two continental plates would produce Which natural event most likely caused the crack in the road? island
Name: Date: 1. The road shown below was suddenly broken by a natural event. 3. The convergence of two continental plates would produce Which natural event most likely caused the crack in the road? island
WEATHER AND CLIMATE practice test
 WEATHER AND CLIMATE practice test Multiple Choice Identify the choice that best completes the statement or answers the question. 1. What role does runoff play in the water cycle? a. It is the process in
WEATHER AND CLIMATE practice test Multiple Choice Identify the choice that best completes the statement or answers the question. 1. What role does runoff play in the water cycle? a. It is the process in
3.1 Measuring Biodiversity
 3.1 Measuring Biodiversity Every year, a news headline reads, New species discovered in. For example, in 2006, scientists discovered 36 new species of fish, corals, and shrimp in the warm ocean waters
3.1 Measuring Biodiversity Every year, a news headline reads, New species discovered in. For example, in 2006, scientists discovered 36 new species of fish, corals, and shrimp in the warm ocean waters
CARBONATE VS SILICICLASTIC DEPOSITIONAL SYSTEMS: A FIELD TRIP GUIDE TO MODERN SEDIMENTARY ENVIRONMENTS AND THE GEOLOGY OF BOCAS DEL TORO, PANAMA
 CARBONATE VS SILICICLASTIC DEPOSITIONAL SYSTEMS: A FIELD TRIP GUIDE TO MODERN SEDIMENTARY ENVIRONMENTS AND THE GEOLOGY OF BOCAS DEL TORO, PANAMA Luis Ignacio Quiroz, Jaime Escobar, Cindy Gonzales, Edward
CARBONATE VS SILICICLASTIC DEPOSITIONAL SYSTEMS: A FIELD TRIP GUIDE TO MODERN SEDIMENTARY ENVIRONMENTS AND THE GEOLOGY OF BOCAS DEL TORO, PANAMA Luis Ignacio Quiroz, Jaime Escobar, Cindy Gonzales, Edward
GEL 113 Historical Geology
 GEL 113 Historical Geology COURSE DESCRIPTION: Prerequisites: GEL 111 Corequisites: None This course covers the geological history of the earth and its life forms. Emphasis is placed on the study of rock
GEL 113 Historical Geology COURSE DESCRIPTION: Prerequisites: GEL 111 Corequisites: None This course covers the geological history of the earth and its life forms. Emphasis is placed on the study of rock
CPO Science and the NGSS
 CPO Science and the NGSS It is no coincidence that the performance expectations in the Next Generation Science Standards (NGSS) are all action-based. The NGSS champion the idea that science content cannot
CPO Science and the NGSS It is no coincidence that the performance expectations in the Next Generation Science Standards (NGSS) are all action-based. The NGSS champion the idea that science content cannot