The neck of Barosaurus: longer, wider and weirder than those of Diplodocus and other diplodocines
|
|
|
- Horatio Gaines
- 7 years ago
- Views:
Transcription
1 The neck of Barosaurus: longer, wider and weirder than those of Diplodocus and other diplodocines Michael P Taylor Corresp., 1, Mathew J Wedel 2 1 Department of Earth Sciences, University of Bristol, Bristol, United Kingdom 2 College of Osteopathic Medicine of the Pacific and College of Podiatric Medicine, Western University of Health Sciences, Pomona, California, United States of America Corresponding Author: Michael P Taylor address: dino@miketaylor.org.uk Barosaurus is a diplodocid sauropod from the Upper Jurassic Morrison Formation of the western United States, and is known for its very long neck. It is closely related to the sympatric Diplodocus, and often thought of as more or less identical except with a longer neck. The holotype YPM 429 includes three and a half posterior cervical vertebrae, somewhat distorted and damaged, which are nevertheless very distinctive and quite different from those of Diplodocus. The cervicals of the better known and more complete referred Barosaurus specimen AMNH 6341 show the same characteristic features as the holotype, though not to the same extent: transversely broad but anteroposteriorly short zygapophyseal facets; prezygapophyses carried on broad, squared-off rami; zygapophyses shifted forward relative to the centrum; diapophyses, parapophyses and neural spines shifted backwards; and broad diapophyseal wings. These features form a single functional complex, enabling great lateral flexibility, but restricting vertical flexibility. This may indicate that Barosaurus used a different feeding style from other sauropods perhaps sweeping out long arcs at ground level. The Morrison Formation contains at least nine diplodocid species in six to eight genera whose relationships are not yet fully understood, but Barosaurus remains distinct from its relatives.
2 1 The neck of Barosaurus: longer, wider and 2 weirder than those of Diplodocus and other 3 diplodocines 4 5 Michael P. Taylor 1, Mathew J. Wedel Department of Earth Sciences, University of Bristol, Bristol, England. 7 dino@miketaylor.org.uk 8 2 College of Osteopathic Medicine of the Pacific and College of Podiatric Medicine, 9 Western University of Health Sciences, Pomona, California, USA. 10 mathew.wedel@gmail.com Abstract 13 Barosaurus is a diplodocid sauropod from the Upper Jurassic Morrison Formation of the 14 western United States, and is known for its very long neck. It is closely related to the 15 sympatric Diplodocus, and often thought of as more or less identical except with a 16 longer neck. The holotype YPM 429 includes three and a half posterior cervical 17 vertebrae, somewhat distorted and damaged, which are nevertheless very distinctive 18 and quite different from those of Diplodocus. The cervicals of the better known and 19 more complete referred Barosaurus specimen AMNH 6341 show the same 20 characteristic features as the holotype, though not to the same extent: transversely 21 broad but anteroposteriorly short zygapophyseal facets; prezygapophyses carried on 22 broad, squared-off rami; zygapophyses shifted forward relative to the centrum; 23 diapophyses, parapophyses and neural spines shifted backwards; and broad 24 diapophyseal wings. These features form a single functional complex, enabling great 25 lateral flexibility, but restricting vertical flexibility. This may indicate that Barosaurus used 26 a different feeding style from other sauropods perhaps sweeping out long arcs at 27 ground level. The Morrison Formation contains at least nine diplodocid species in six to 28 eight genera whose relationships are not yet fully understood, but Barosaurus remains 29 distinct from its relatives Keywords: dinosaur, sauropod, Barosaurus, Diplodocus, neck, cervical vertebrae 32
3 33 Introduction 34 Barosaurus is an iconic genus of sauropod dinosaur, a diplodocine diplodocid from the 35 Morrison Formation of Late Jurassic western United States (Marsh 1890, Lull 1919). 36 Even among sauropods, its neck is proportionally and absolutely very long at about m (Wedel 2007: ) the same length as the neck of the much bulkier African 38 brachiosaur Giraffatitan, and three and a half times as long as that of the world-record 39 giraffe (Toon and Toon 2003:399). As such, it is a staple in popular dinosaur books (e.g. 40 Bartram et al. 1983, Lindsay 1992, Lambert 2000). A mounted cast of a Barosaurus 41 skeleton, AMNH 6341, dominates the entrance hall of the American Museum of Natural 42 History, dwarfing the adjacent skeleton of the predatory dinosaur Allosaurus (Figure 1). 43 A recently rediscovered Barosaurus skeleton, ROM 3670, has been mounted at the 44 Royal Ontario Museum where it provides the centrepiece of the dinosaur gallery. Other 45 specimens are known, but are less complete and in many cases juvenile, so less 46 informative. The complex history of the genus and its principal specimens is helpfully 47 summarised by McIntosh (2005:40 43). 48 Barosaurus is sometimes thought of as merely Diplodocus with a longer neck. In fact, 49 the cervical vertebrae of Barosaurus are not merely elongated versions of those of its 50 relative, but morphologically very distinct. Here, we reconsider the cervicals of the 51 holotype specimen, compare them with those of the referred AMNH specimen, 52 recognise distinctive features of the Barosaurus neck, and consider their functional 53 implications. 54 Institutional abbreviations 55 AMNH American Museum of Natural History, New York (USA). 56 ANS Academy of Natural Sciences, Philadelphia (USA). 57 MB Museum für Naturkunde Berlin, Berlin (Germany). 58 NSMT National Science Museum, Tokyo (Japan). 59 ROM Royal Ontario Museum, Toronto (Canada). 60 SMA Sauriermuseum Aathal (Switzerland). 61 YPM Yale Peabody Museum, New Haven (USA). 62 Description 63 Comparison of Barosaurus and Diplodocus 64 As noted by McIntosh (1990: ), Barosaurus resembles Diplodocus in most 65 aspects of its anatomy: 66 The genus [Barosaurus] is very closely related to Diplodocus, and the limb 67 bones are so similar as to be indistinguishable. Barosaurus differs from 68 Diplodocus in its enormously elongated cervical vertebrae, which are
4 69 relatively 33 percent longer than those of the latter. [ ] The enormously 70 elongated cervicals are generally similar to those of Diplodocus if the latter 71 were stretched. 72 As such, Barosaurus is invariably depicted as virtually identical to Diplodocus except for 73 an elongated neck as for example in the skeletal reconstructions of Paul (2000:406). It 74 has even been tentatively suggested by Senter (2006:46) that Diplodocus and 75 Barosaurus might be sexual dimorphs, with the longer neck of the latter marking it out 76 as the more flamboyant male. Such possibilities are lent credibility by the close 77 phylogenetic position of the two taxa: every phylogenetic analysis that includes both 78 genera has recovered them as sister taxa, including the sauropod phylogeny in The 79 Dinosauria, 2nd edition (Upchurch et al. 2004: fig. 13:18) and the recent diplodocoid 80 phylogenetic analyses of Whitlock (2011: fig. 7) and Mannion et al. (2011: fig. 10). 81 What is more, autapomorphies of the Barosaurus neck have been surprisingly hard to 82 find in the literature. The phylogenetic analysis of Wilson (2002) gives three 83 autapomorphies for Barosaurus, but two are in the dorsal vertebrae and one in the 84 caudals. The analysis of Upchurch et al. (2004) also finds two autapomorphies in the 85 dorsals and one in the caudals, though these are different from those of Wilson (2002). 86 In the analysis of Taylor et al. (2011b), based on that of Harris (2006b), four 87 autapomorphies were found, but three were again in the dorsal vertebrae and one in the 88 ischium. No autapomorphies of Barosaurus are given by Whitlock (2011) or Mannion et 89 al. (2011). Similarly, McIntosh (2005:39) gave a differential diagnosis separating 90 Barosaurus from Diplodocus, but the only cervical characters listed are the presumed 91 increase in cervical count, and elongation of the vertebrae. Consequently, and 92 surprisingly, none of these analyses reported any autapomorphies in the neck of 93 Barosaurus, its most distinctive feature. This indicates that additional characters, 94 discussed below, should be added to future analyses. 95 Despite the failure of modern studies to identify differences between the cervicals of the 96 two diplodocines, Lull (1919:20), in his classic descriptive monograph of Barosaurus, 97 was cautious regarding the relationship between these genera: 98 The preserved elements compare most nearly with those of Diplodocus, but 99 differ remarkably in certain proportions. These resemblances may have been 100 in part convergence and merely similar mechanical adjustments of bony 101 tissue to meet similarly disposed strains and stresses, and as such imply no 102 close relationship. 103 The post-cervical skeletons of Diplodocus and Barosaurus are indeed very similar, 104 although the latter has only nine rather than ten dorsal vertebra, having apparently 105 recruited the anteriormost dorsal into its neck (McIntosh 2005:44 45), and has a shorter 106 tail (McIntosh 2005:57). However, the widely assumed similarity of the cervical 107 vertebrae between the diplodocines is based almost entirely on lateral views (Figure 2). 108 This is understandable, as the cervical columns of both Diplodocus and Barosaurus 109 have been illustrated in detail in lateral view the former as both drawings and 110 photographs by Hatcher (1901: plates III and IV), the latter as photographs only by 111 McIntosh (2005: fig. 2.1). However, the cervical vertebrae of Diplodocus have been 112 illustrated in anterior and posterior views only by relatively uninformative photographs
5 113 (Hatcher 1901: plates V and VI), and only two cervicals of Barosaurus (C8 and 13) have 114 been illustrated in anterior or posterior views (McIntosh 2005: fig. 2.2). Worst of all, the 115 important dorsal view is completely unpublished for Diplodocus, and published for only 116 a single vertebra in the Barosaurus holotype (Lull 1919: plate II: part 3). The vertebra 117 illustrated by Lull is part of the holotype specimen YPM 429, designated by him as 118 vertebra R and considered by him to be the most posterior cervical. It is extremely 119 distinctive and superficially very different from those of Diplodocus (Figure 3). 120 Although the cervical series of Diplodocus has not been illustrated in dorsal view, those 121 of three other diplodocids have: Apatosaurus ajax (Upchurch et al. 2005: plate I), 122 Suuwassea (Harris 2006a: figs. 4 9; note that some recent phylogenies place this at the 123 base of Dicraeosauridae, the sister group to Diplodocidae), and most helpfully 124 Kaatedocus (Tschopp and Mateus 2012: unnumbered supplementary figures). The 125 Kaatedocus figures of Tschopp and Mateus are full colour, high resolution photographs 126 of all fourteen preserved cervical vertebrae in five cardinal directions, and so provide an 127 invaluable comparative resource especially as Kaatedocus is probably a diplodocine, 128 and so more closely related to Diplodocus and Barosaurus than Apatosaurus and 129 Suuwassea are (Taylor and Naish 2005: table 1). 130 Comparison of the posterior cervicals of these genera in dorsal view (Figure 4) appears 131 to show that Barosaurus is dramatically different from the others: for example, it has 132 very broad prezygapophyseal rami that are squared off anteriorly, wide wings that 133 sweep back to posteriorly placed diapophyses before cutting back in towards the 134 centrum, and an apparently unsplit neural spine at the junction of an X shape formed 135 by the spinoprezygapophyseal and spinopostzygapophyseal laminae. 136 The neck of the Yale Barosaurus 137 The genus Barosaurus has only one species, B. lentus, and the holotype specimen, 138 YPM 429, is held at the Yale Peabody Museum. As noted by Lull, this specimen 139 contains material from four posterior cervical vertebrae, which he arbitrarily designated 140 as vertebrae Q, R, S and T. Of these, vertebra T is too incomplete to be informative, but 141 the other three are all informative. Measurements are given in Table 1. Table 1. Measurement of cervical vertebrae in YPM 492, the Barosaurus lentus holotype. Measurements taken from Lull (1919) are suffixed L ; measurements from photographs are marked P. Width across parapophyses of vertebra Q based on reconstruction of how the undamaged element would have been. All measurements are in mm. Vertebra Vertebra R Vertebra Q Vertebra S Serial position C?15 C?13 C?12 Total length 960 L L Total height 560 L Centrum length L Condyle height L Condyle width L 220 L Condyle height:width ratio Cotyle height L Cotyle width L
6 Vertebra Vertebra R Vertebra Q Vertebra S Cotyle height:width ratio Width across prezygapophyses 620 Width across diapophyses Width across parapophyses 410 P 330 P 200 P Left prezygapophyseal ramus width (anterior P end) Right prezygapophyseal ramus width (anterior 240 end) Left prezygapophyseal facet width 126 P Right prezygapophyseal facet width Vertebra R 143 Because Lull's vertebra R differs significantly from all other diplodocid vertebrae in 144 dorsal, lateral and anterior views (compare Figure 3 with Figure 2 and with Hatcher : plate V), it is appropriate to consider whether it really is part of the same 146 individual as the rest of YPM 429; and, if so, whether YPM 429 is really a diplodocid at 147 all, and whether AMNH 6341 and other classic Barosaurus individuals have been 148 incorrectly referred. 149 Vertebra R resembles the cervicals of brachiosaurids and other basal titanosauriforms 150 rather than those of diplodocids in the following respects: 151 Its neural arch and spine are much less tall relative to total length, as seen in 152 Giraffatitan (Janensch 1950: figs ), Brachiosaurus sp. (BYU 12866, Wedel : fig. 7.2) and especially Sauroposeidon (Wedel et al. 2000a, b). 154 The articular surfaces of its centrum are anterodorsally inclined, as in the 155 Giraffatitan lectotype specimen MB.R.2180 (previously known as HMN SI; 156 Janensch 1950: figs ). 157 The entire neural arch is shifted forward on its centrum, so that the 158 prezygapophyses greatly overhang the anteriormost part of the centrum, and the 159 postzygapophyses (which are broken off) must have been located corresponding 160 forward from the posterior rim of the centrum, as in Giraffatitan, Brachiosaurus, 161 and Sauroposeidon. 162 The parapophyses are located more posteriorly than the diapophyses, so that a 163 line joining them is inclined anterodorsally rather than posterodorsally, as in at 164 least some vertebrae of Giraffatitan and Brachiosaurus. 165 The neural spine appears unsplit, rising to a low peak that is buttressed from the 166 four diagonals by zygapophyseal laminae (compare with Figure 5). 167 On the other hand, vertebra R also has some significant dissimilarities to brachiosaur 168 cervicals: 169 Its neural spine appears proportionally lower than in any known posterior 170 vertebra of a sauropod, its condition being perhaps most closely approached by 171 C6 of MB.R.2180 (Janensch 1950: fig. 26).
7 172 The vertebra is very much more proportionally broad than in brachiosaurs, 173 resembling in this respect (though not in others) the vertebrae of some 174 titanosaurs such as Puertasaurus (Novas et al. 2005: fig. 1). 175 The prezygapophyseal rami are extraordinarily broad, whereas those of 176 brachiosaurs are drawn forward almost to a point, where they bear small oval 177 facets (Figure 5). 178 The distinctiveness of vertebra R raises important questions about the Barosaurus 179 holotype YPM 429. Does it belong to an animal very different from the classical 180 conception of Barosaurus, which is derived primarily from AMNH 6341? Or perhaps 181 YPM 429 as a whole is similar to the AMNH specimen, but vertebra R is part of a 182 different animal that was inadvertently referred to the same specimen? The latter seems 183 unlikely, as the quarry map (Lull 1919: fig. 2) shows it closely associated with the other 184 three cervical vertebrae, and surrounded on three sides by other elements belonging to 185 the specimen. The solution to the mystery of vertebra R, then, is to be found in the other 186 cervical vertebrae that are part of YPM Vertebra Q 188 Vertebra Q is similar in size to vertebra R: it is 2% longer overall (980 vs. 960 mm), 189 though its centrum is fully 22% longer (820 vs. 670 mm). But it is proportionally 190 narrower: only 80% as broad across the diapophyses (580 vs. 720 mm). Helpfully, it is 191 preserved upside down in its jacket, and so different portions of the vertebra are 192 available for study (Figure 6). Although it is less in overall breadth than vertebra R, it 193 shares some important features that corroborate Lull's assignment of both to the same 194 individual. Most importantly, the left prezygapophysis is preserved and undistorted, and 195 is very broad as in vertebra R. (The prezygapophyses of vertebra R seem to be broader 196 still, but distortion and reconstruction make it difficult to be certain of their true width.) 197 Vertebra Q also has wing-like prezygadiapophyseal laminae that are swept back like 198 those of vertebra R. And, contra Lull (1919:14), the diapophyses of vertebra Q are 199 positioned more anteriorly than its parapophyses, as in vertebra R (Figure 6: top part). 200 Assuming that the two vertebrae do belong to the same individual, vertebra Q adds 201 important information. Its postzygapophyses are unbroken: they sweep out 202 posterolaterally and upwards from behind the diapophyseal wings and appear triangular 203 in posterior view. The postzygapophyseal facets are difficult to discern precisely, but 204 seem to be very broad, extending almost all the way to the lateral edges of the rami that 205 bear them, and so matching the broad prezygapophyseal facet that is apparent in 206 anterior view (Figure 6: left part). 207 Most significantly, when viewed in left ventrolateral aspect, vertebra Q can be seen to 208 bear a left metapophysis, broadly similar in shape to what would be expected in a 209 diplodocid: flat and somewhat laminated, anteroposteriorly longer than tall, and with 210 distinct anterodorsal and posterodorsal corners. The right metapophysis is either lost or 211 embedded in the jacket.
8 212 Vertebra S 213 Vertebra S is the longest of the preserved vertebrae: it is 13% longer than vertebra Q in 214 centrum length (930 vs. 820 mm) and 4% longer in total preserved length (1020 vs mm). Its preservation is very different from that of vertebrae R and Q. While those 216 vertebrae present their dorsal and ventral faces respectively, and have undergone some 217 dorsoventral crushing, vertebra S lies on its left side in its jacket so that the right lateral 218 view is presented (Figure 7), and it appears to have been crushed transversely. Its 219 cotyle height is 124% of its width compared with 53% and 51% for R and Q 220 respectively; and the preserved width across parapophyses is only 200 mm compared 221 with 410 and 330 for R and Q (Figure 8). 222 Vertebra S provides the clearest evidence of bifid neural spines in YPM 429, as both 223 metapophyses are preserved. These are apparent in dorsal view. The 224 intermetapophyseal cleft is shallow, only about 75 mm deep. In Barosaurus the 225 bifurcation of cervical neural spines starts farther back along the neck than it does in 226 Diplodocus, and as far back as C13 in AMNH 6341 only a shallow cleft is present 227 (McIntosh 2005: fig. 2.3A). 228 Both prezygapophyseal rami are present but incomplete. The better preserved left 229 ramus indicates that despite its much lesser overall broadness, this vertebra had broad 230 prezygapophyses similar in character if not in degree to those of vertebrae R and Q. 231 The right ramus is more distorted, the spinoprezygapophyseal lamina having been 232 displaced in a lateral kink. 233 The right postzygapophysis is intact. As with vertebra Q, the facet is broad, and is 234 supported by a wide ramus that is strongly triangular in dorsal or ventral view. 235 The cortex of vertebra S has eroded away from the condyle, revealing a camellate 236 internal structure of many small, irregular pneumatic cells. Similar structure is also 237 visible, though less clearly, in the broken condyle of vertebra Q. Lull (1919:11) noted 238 that the pneumatic fossae in the lateral faces of the centra of Barosaurus cervicals are 239 consistently smaller than those in equivalent vertebrae of Diplodocus, though no less 240 deep: this external morphology is consistent with that of titanosaur presacrals, which 241 also have camellate to somphospondylous internal structure, suggesting that the 242 internal and external structures are functionally correlated. 243 Association of the cervical vertebrae 244 In light of their similar general morphology overall broadness, prezygapophyses 245 extending well forwards of their centra, very broad prezygapophyseal rami and facets 246 it is reasonable to assume that vertebrae R and Q belong to the same individual. But 247 vertebra S poses a problem: it is much narrower than the other two vertebrae, and 248 correspondingly taller; and the preserved portions of its prezygapophyses hardly 249 overhang its centrum at all. 250 However most of these differences can be explained by the different preservation of the 251 three vertebrae. The orientation of the elements in their jackets alone is sufficient to 252 suggest that only vertebra S was found on its side. If it suffered a moderate amount of 253 transverse crushing and the other two were affected by dorsoventral crushing, then all
9 254 three could have approached an intermediate morphology when fully intact. 255 The lack of prezygapophyseal overhang in vertebra S can also be explained: an 256 additional bony plate is preserved, above the condyle but unattached (Figure 7: inset). It 257 resembles the flat surfaces of the prezygapophyseal rami of the other vertebrae. It must 258 represent the anterior portion of one of the rami, broken downwards and inwards. So 259 most likely vertebra S did have overhanging prezygapophyses, and therefore had a 260 greater total length when intact. 261 Other features suggest a relationship between vertebrae Q and S. They share the 262 distinctive triangular shape of the postzygapophyses as seen from below (also seen in 263 Kaatedocus), and both have small fossae just below the tip of the metapophysis (also 264 present in Dinheirosaurus). 265 There is little to tie vertebra R directly to S, but Q is a helpful intermediate both in 266 preservation and possibly in serial position which is evidently similar to both, and so 267 ties them together. 268 Interesting differences among the vertebrae remain even after accounting for 269 taphonomic deformation. First, in vertebrae R and Q, but not in S, the diapophyses are 270 more anteriorly positioned than the parapophyses, and this remains true even when 271 vertebra R is corrected for shearing. This can only be interpreted as serial variation 272 between individual vertebrae. Something similar is seen in Hatcher's (1901: plate III) 273 illustration of the cervicals of Diplodocus carnegii, in which the diapophysis of C12 is 274 directly above the parapophysis whereas it is more posterior in all the other cervicals. 275 Second, Lull's (1919: 11) description states, and our observations confirm (Figure 7), 276 that a small midline keel is present on the ventral surface of Vertebra S. Vertebrae R 277 and Q have no trace of a keel. The presence of a ventral keel in the cervical vertebrae 278 is a primitive character for sauropods, and keels are present in Barapasaurus, 279 Shunosaurus, Patagosaurus, Omeisaurus, Mamenchisaurus, and Phuwiangosaurus 280 (Upchurch 1998). Among diplodocoids, ventral keels are present in the cervical 281 vertebrae of Dicraeosaurus (Upchurch 1998 and pers. obs.) and they are variably 282 present in Haplocanthosaurus (Wedel and Sanders 2002: 2). More delicate ventral 283 ridges appear in some cervical vertebrae at BYU that are probably referable to 284 Barosaurus (pers. obs.) and in certain privately held specimens. However, the number 285 and nature of these ridges in diplodocids is highly variable, even between different 286 vertebrae of the same individual. These features may or may not be homologous with 287 the true ventral ridges of basal sauropods and dicraeosaurs, but are probably not 288 diagnostic at the generic level. 289 Reconstructions of the cervical vertebrae 290 Vertebra R is probably the most distorted, having evidently undergone not only crushing 291 but also shearing, with the dorsal part of the vertebra shifted anteriorly (Lull 1919:14) 292 which has exaggerated the already substantial prezygapophyseal overhang and shifted 293 the diapophyses further forward of the parapophyses than they would have been. 294 In addition, both postzygapophyses are missing. These can be reconstructed after 295 those of vertebra Q, but perhaps splaying further laterally than in Q to correspond with
10 296 R's broader prezygapophyses. 297 Because the spinoprezygapophyseal and spinopostzygapophyseal laminae converge to 298 a low point, with ossified ligament attached to its posterior aspect, we were initially 299 inclined to perceive this as the summit of an unusually low neural spine. However, the 300 edges of the laminae do not preserve any finished bone, instead being broken in some 301 places and restored with plaster in others. In light of the clearly bifid spine for vertebra 302 S, and of the single preserved metapophysis of vertebra Q, we now accept the 303 interpretation of Lull (1919:14), that vertebra R in life bore plate-like metapophyses that 304 rose well above the level of the highest preserved point, and the remaining parts of the 305 spinopostzygapophyseal and spinopostzygapophyseal laminae represent the bases of 306 these lost metapophyses (Figure 9). 307 We initially suspected that the prezygapophyseal rami were broken off and would have 308 extended yet further anteriorly in life. This was based on three things: the assumption 309 that they could not have been so broad at their extremity; the folded profile of the rami in 310 anterior aspect, which could not bear functional articular facets; and the lack of 311 perceptible finished bone along much of the anterior margin. However, all of these 312 points now seem flawed: the broadness of the rami is a genuine osteological feature, 313 corroborated by the similar (though less extreme) morphology in vertebra Q; the rami 314 appear folded because they have indeed been folded by crushing, and would have 315 been straighter in life; and the paucity of good bone along the anterior margin is due to 316 over-enthusiastic restoration work and the liberal application of plaster. So we now feel 317 that the complete zygapophyseal rami are preserved, though badly damaged. 318 Vertebra Q seems to be less distorted, but it has undergone a complex crushing along a 319 diagonal axis along with some twisting. Although it does not lie in a true upside-down 320 position in its jacket, the ventral aspect shows that most of the element is intact apart 321 from the right anterolateral portion including the right prezygapophysis, parapophysis, 322 and much of the condyle. These can mostly be reconstructed by mirroring from the 323 better left side. 324 While well preserved in most respects, vertebra S is missing its entire diapophyseal 325 wing and the anterior tips of both prezygapophyses. As a result it is superficially very 326 different from the other two cervicals. However, the anterior part of one 327 prezygapophysis is present, out of position above the centrum; and the missing parts 328 can be tentatively reconstructed by reference to vertebra Q. 329 Serial position of the cervical vertebrae 330 Lull (1919:11 15) considered these three vertebrae, together with the fragmentary and 331 uninformative vertebra T, to be the four most posterior cervicals C12 15 of his usage, 332 since he thought Barosaurus, like Diplodocus, had 15 cervicals. He placed them in the 333 sequence S, Q, T, R from front to back, determined in part by the circumference of the 334 posterior articular face of the centrum (p. 11), but he did not mention any other criteria. 335 Lull's relative positions for the three adequately preserved vertebrae are corroborated 336 by their progressively decreasing length and increasing broadness across the 337 parapophyses (Table 1; Figure 8): compare with Janensch's (1950: fig. 50) illustration of 338 the ventral view of vertebrae C10 C13 and D1 D2 of Giraffatitan; and with Upchurch et
11 339 al.'s (2005: plate 1) illustration of the dorsal views of cervical vertebrae of Apatosaurus. 340 Even allowing for some transverse crushing of vertebra S, it must have been narrower 341 than vertebra Q when intact. The featureless ventral surface of vertebra R also 342 corroborates its position as the most posterior of the preserved cervicals, as this 343 condition is often seen in posterior cervicals and in dorsals. 344 The absolute positions of the vertebrae are harder to judge. The centra appear rather 345 elongate to be the most posterior cervicals as suggested by Lull: specifically, vertebra 346 R, probably the most posterior of the three, is somewhat longer than the last cervical of 347 the AMNH specimen (960 vs. 750 mm). However, the vertebrae are evidently close to 348 the back of the neck. They were found in association with an anterior dorsal (considered 349 D1 by Lull, but reassigned as D2 by McIntosh 2005:48), which does suggest that there 350 were probably not many intervening vertebrae. Accordingly, we tentatively consider 351 these to be three of the four cervicals before the last, i.e. C12 C15. Another possibility 352 is that the broken vertebra T of which only the most posterior part remains was the 353 last cervical, C16, and S, Q and R are C13 C15. But Lull (1919:14 15) points out that 354 the cotyle circumference of vertebra T is intermediate between that of vertebrae Q and 355 R. So our preferred interpretation is that S is C12, Q is C13, T is C14, R is C15, and 356 C16 and D1 are missing. 357 The neck of the AMNH Barosaurus 358 Comparisons between the referred Barosaurus individual AMNH 6341 and the holotype 359 YPM 429 have been hampered by the lack of published illustrations of the AMNH 360 material. McIntosh (2005: fig. 2.1) illustrated the preserved cervicals (C8 C16) but only 361 with small, poorly reproduced monochrome photographs of the left lateral view. Two of 362 the vertebrae are also illustrated in anterior or posterior view C8 in fig. 2.2A and C in fig. 2.3A, but the remainder are not. None are illustrated in dorsal view. 364 Unfortunately these vertebrae are now inaccessible for study: they are on display in the 365 Hall of Saurischian Dinosaurs at the American Museum of Natural History, but 366 inconveniently located underneath a glass walkway which is scuffed by the feet of 367 visitors. As a result, photography is very difficult. Nevertheless, because there are 368 currently no published dorsal-view illustrations, we have made our best effort to capture 369 the vertebrae from above and to clean the resulting images (Figures 10, 11). 370 It is now apparent that the penultimate cervical of the AMNH specimen bears important 371 similarities, not previously apparent, to vertebra R (Figure 12): 372 While not as broad as those of vertebra R, the prezygapophyseal rami of the 373 AMNH vertebra are much broader and squarer in dorsal view than in other 374 sauropods: compare with Figure The prezygadiapophyseal laminae form broad horizontal wings, which sweep 376 inwards towards the centrum behind the diapophysis. 377 The bases of the metapophyses converge at the midline and form an X 378 composed of the spinozygapophyseal laminae, as in vertebra R, corroborating 379 the interpretation that this vertebra originally bore metapophyses that have since 380 been lost.
12 381 These similarities suffice to confirm the referral of the AMNH material to Barosaurus: the 382 remaining differences in proportion between vertebra R and the AMNH cervical can 383 mostly be understood as the result of individual variation or differences in preservation. 384 The similarities between C15 of the AMNH material and vertebra R of the YPM series 385 lend credence to the idea that the latter series really does represent C12 15, as 386 outlined above. 387 In some of the dorsal-view photographs of the AMNH cervicals, the prezygapophyseal 388 facets can be discerned, verifying that they are both transversely broad, occupying 389 almost the whole width of the rami, and anteroposteriorly short. It is not possible to 390 determine prezygapophyseal facet extent directly from the Yale material due to poor 391 preservation and over-enthusiastic reconstruction of this area in vertebra R, but it must 392 be assumed to resemble the condition in the AMNH material. 393 Discussion 394 Fusion of vertebral elements 395 The diapophyses and left parapophysis of vertebra Q preserve articular surfaces, 396 indicating that the cervical ribs were unfused in this individual despite its great size 397 surprisingly, as McIntosh (2005:48) says that in the similarly sized or slightly smaller 398 AMNH 6341 The cervical ribs are firmly coalesced to all the cervicals. Parapophyseal 399 and diapophyseal facets are also present in vertebra R, though poorly preserved and 400 difficult to interpret. They are lost in vertebra S. 401 Hatcher (1901: plate III) shows vertebrae in C6 C15 of Diplodocus carnegii CM 84 with 402 their ribs fused to them (though broken in C10). Therefore, either YPM 429 was 403 considerably less mature than CM 84, despite being of comparable size, or Barosaurus 404 and Diplodocus did not follow the same ontogenetic trajectory of fusions. This 405 represents another example of the increasingly recognised inconsistency in the timing 406 of fusions in sauropod ontogeny (Wedel and Taylor 2013: table 1; Hone et al. 2016). 407 The discrepancies between ontogenetic progression in YPM 429, AMNH 6341 and CM are further evidence that lumping multiple taxa together in analyses of ontogenetic 409 change (e.g. Woodruff and Fowler 2012) is unwise. 410 Functional implications of Barosaurus neck anatomy 411 Functional implications follow from the unique anatomy of the Yale Barosaurus material. 412 The short anteroposterior extent of the zygapophyseal facets together with the anterior 413 displacement of the zygapophyseal articulations relative to those of the centra suggest 414 that the neck may have been limited in vertical flexibility. On the other hand, the 415 extreme transverse width of the facets seems to indicate an unusual degree of lateral 416 flexibility. Lull (1919:13) recognised the latter, but did not comment on the former. 417 The broad diapophyseal wings of Barosaurus, the posterior migration of the 418 diapophyses and parapophyses, and the anterior extension of the zygapophyses would 419 also have had implications for lateral movement of the neck. The broadness of the 420 wings shifted the lateral muscles away from the midline, allowing them to act with
13 421 greater mechanical advantage. Each of the other changes contributed to extending the 422 length of the ansae costotransversariae, or cervical rib loops, which provided the 423 attachment area for the long lateral flexors. Assuming that these muscles were laid out 424 as they are in birds (see Wedel and Sanders 2002), the Mm. cervicalis ascendens 425 originated on the prezygadiapophyseal laminae and inserted on the epipophyses of 426 more anterior vertebrae; and the Mm. flexor colli lateralis originated on the anterior 427 aspect of the rib, including its anterior projection. Both these areas were proportionally 428 larger in Barosaurus than in other diplodocines (Figure 13). 429 The parapophyses and diapophyses migrate progressively backwards in the posterior 430 vertebrae of Diplodocus (Hatcher 1901: plate III) as well as in Barosaurus; but the 431 migration seems to begin more anteriorly in Barosaurus (Lull 1919:12) and reaches its 432 extreme in vertebra R, where the prezygadiapophyseal laminae run half the entire 433 length of the vertebra. By contrast the diapophyses and parapophyses do not migrate 434 backwards in the posterior cervicals of Kaatedocus (Tschopp and Mateus: unnumbered 435 supplementary figures), and together with the mediolateral narrowness of the 436 Kaatedocus cervicals this may indicate that lateral neck motion was less important in 437 this taxon (although this narrowness is increased by transverse compression in SMA ). 439 So the broad zygapophyseal facets, diapophyseal wings and posterior migration of the 440 cervical rib loop in Barosaurus are all aspects of a single functional complex related to 441 lateral neck movement: the facets enable lateral flexibility and the other features provide 442 both expanded attachment area and mechanical advantage to the muscles that produce 443 it. Not only was the neck of Barosaurus absolutely and proportionally long even among 444 sauropods, it was also uniquely adapted for lateral sweeping: no other sauropod shows 445 the same degree of development of the relevant characters. 446 Like the diapophyses and parapophyses, the neural spine is also located more 447 posteriorly in Barosaurus than in other diplodocids. Two groups of muscles are inferred 448 to originate on the spine (Wedel and Sanders 2002): the Mm. interspinales, which insert 449 on the posterior aspect of the next spine, and so are invariably the length of a single 450 vertebra; and the Mm. longus colli dorsalis, which span many vertebrae, so that 451 posterior displacement of the origin within a single vertebra would have very little 452 mechanical effect. We are therefore unable to determine what purpose if any the 453 posterior location of the neural spine served, although we note that the apex of the 454 neural spine is also located quite far back in posterior cervicals of Giraffatitan and 455 Sauroposeidon (Janensch 1950, Wedel et al. 2000a, b), so this character seems to be 456 correlated with neck elongation. 457 Behavioural implications 458 How did Barosaurus use its unique neck? Martin (1987) proposed, and Stevens and 459 Parrish (1999) and Ruxton and Wilkinson (2011) developed, a vacuum-cleaner 460 feeding hypothesis for sauropods: that they spent much of their time standing stationary 461 and feeding at ground level, sweeping out a broad area with their long necks. Although 462 we have been sceptical that this feeding model was common for sauropods (Taylor et 463 al. 2009, 2011a), we recognise that Barosaurus, with its limited dorsoventral flexibility at
14 464 the base of the neck and its suite of lateral-sweep adaptations, is a prime candidate for 465 such behaviour. 466 Most work on niche partitioning among Morrison sauropods has focused on their jaws 467 and teeth (e.g. Barrett and Upchurch 1994, Fiorillo 1998, Button et al. 2013). Browsing 468 height has also been mentioned in connection with niche partitioning (e.g. Paul 1998). 469 However, beyond the general discussion of Stevens and Parrish (2005a, 2005b), the 470 role of neck behaviour has so far been little studied. But sauropod heads were, to a first 471 approximation, simple food scoops; and their necks were almost certainly adapted 472 primarily for food gathering (Sander et al. 2010, Taylor et al. 2011a). Given these 473 baseline similarities it seems likely that different sauropod taxa were using their necks in 474 different ways. 475 Morphological trends in the necks of diplodocines 476 Barosaurus is not just a stretch-limo remix of Diplodocus. Not only is the morphology of 477 its neck different in functionally significant ways, but as noted by Lull (1919:34 36), the 478 pubis of the type specimen is significantly larger than that of the similar-sized 479 Diplodocus carnegii. (Lull also described additional appendicular elements of the 480 Barosaurus holotype, all larger or more robust than their counterparts in Diplodocus. But 481 McIntosh (2005:40 41) says that these are from a different site, a fact that Lull 482 singularly fails to record, and says that there is no reason to believe they belong to YPM , or to Barosaurus at all. The pubis is the only appendicular element shown in the 484 quarry map of Lull 1919: fig. 2, and the only one that can be confidently associated with 485 the holotype.) 486 The cervical vertebrae of the Morrison-Formation diplodocines Diplodocus, Kaatedocus, 487 AMNH Barosaurus and Yale Barosaurus seem to form a continuum: each stage in the 488 sequence has more elongate centra, its zygapophyses shifted further forward with 489 respect to the centrum, its neural spine further back, its diapophyseal wings broader 490 and its prezygapophyseal rami broader and more squared off in dorsal view. 491 Differences remain between the Yale and AMNH Barosaurus cervical material, even 492 when allowing for differences in preservation. The trend towards broadening the 493 zygapophyses and the diapophyseal wings is taken to an extreme in the Yale material. 494 This is best seen by comparing vertebra Q, which we tentatively identify as C13, with 495 C12 of the AMNH specimen. When scaled to the same total length, the Yale vertebra is % wider across the diapophyses and 95% wider across the postzygapophyses. This 497 could possibly indicate that the two specimens represent different species; it could be 498 sexual dimorphism, with the male exhibiting a flamboyant neck; or it might simply be 499 individual variation. 500 It is also possible that the differences between the necks of the Yale and AMNH 501 Barosaurus specimens represent changes through evolutionary time. Unfortunately, this 502 possibility cannot be meaningfully evaluated, as the stratigraphic context of the YPM 503 specimen is not well constrained. As explained by Foster (2013), limited exposure of the 504 Morrison Formation in the region of the quarry makes it unclear whether the YPM 505 specimen is older or more recent than the AMNH one.
15 506 Diplodocid diversity in the Morrison Formation 507 With the recent addition of Kaatedocus to the roster, the diversity of diplodocids in the 508 Morrison Formation has become yet more impressive: as well as the diplodocines 509 Diplodocus, Galeamopus (probably two species), Barosaurus and Kaatedocus, there 510 are at least four species of Apatosaurus (Upchurch et al. 2005: fig 15), Supersaurus, 511 and possibly Eobrontosaurus, which awaits restudy. Other diplodocoids are also 512 present in the Morrison Formation: Suuwassea, which is now thought to be a 513 dicraeosaurid (Whitlock 2011); probably Amphicoelias (Whitlock 2011, Mannion et al ); and possibly Haplocanthosaurus, which was recovered as a basal diplodocoid in 515 the analyses of Wilson (2002), Whitlock (2011) and Mannion et al. (2011). 516 It's interesting that of the 13 or so diplodocoid species currently known from the 517 Morrison Formation, 10 are diplodocids. As noted by Taylor (2006), the clade 518 Diplodocidae was limited in time and space: more diplodocids are known from the 519 Morrison Formation than from the rest of the global Mesozoic put together. Yet in the 520 one time and place when Diplodocidae flourished, its diversity was much greater than 521 that of other sauropod groups. By comparison the other diplodocoid clades, 522 Rebbachisauridae and Dicraeosauridae, were less speciose at any given time but 523 longer lived. 524 The very high diversity of sauropods in the Morrison Formation gives us a picture of an 525 amazing ecosystem positively abundant with numerous species of giant animals bigger 526 than anything alive on land today. It could be argued that the extremity of such an 527 ecosystem constitutes evidence that Morrison sauropods are oversplit. But this 528 argument from incredulity would be mistaken. We must assess taxonomy on its own 529 grounds, based on what the fossil morphology tells us; only then can we determine what 530 the resulting species roll-call tells us about the ecosystem. Mesozoic ecosystem were 531 simply not like modern ones (see e.g. O Gorman and Hone 2012), and sauropod 532 possessed a suite of key adaptations that have not been combined in any other clade of 533 organisms. We must resist the insidious temptation to assume that what we would have 534 seen in the Late Jurassic is somehow analogous to what we see today on the 535 Serengeti. 536 Trends in sauropod descriptive monographs 537 The classic descriptive monographs on sauropods remain influential and useful (e.g. 538 Hatcher 1901 on Diplodocus, Lull 1919 on Barosaurus, Gilmore 1936 on Apatosaurus, 539 Janensch 1950 and other papers on Giraffatitan). However, they are showing their age, 540 and due for revision. They were mostly written at a time when only a tiny fraction of 541 presently recognised sauropod diversity was known, and without phylogenetic context. 542 The illustrations in these monographs, while aesthetically beautiful, are often less 543 scientifically uninformative than those of modern descriptions, depicting elements in 544 only one or two orientations, invariably in monochrome, and often at small sizes. 545 With the increasing accessibility of digital photography and online publishing, fossils 546 should now be routinely illustrated from as many of the cardinal directions as possible, 547 in full colour and at high resolution. The excellent multi-view photographs of the 548 Kaatedocus cervicals provided with the description of Tschopp and Mateus (2012)
16 549 demonstrate what is now possible, and set a new bar for descriptive illustration though 550 it is unfortunate that they are not part of the main paper, but relegated to second-class 551 status as unnumbered supplementary figures. 552 Conclusions 553 Barosaurus is a valid genus of diplodocine sauropod, and the specimen AMNH from which it is principally known is closely related to the holotype YPM Barosaurus is distinguished from all other sauropods by the nature of its cervical 556 vertebrae. These bear uniquely broad and anteroposteriorly short prezygapophyseal 557 facets on uniquely broad prezygapophyseal rami, which merge into broad, wing-like, 558 horizontal prezygadiapophyseal laminae. This suggests that the neck of Barosaurus 559 was mechanically optimised for wide, sweeping horizontal movements, but may have 560 been less mobile vertically. 561 Acknowledgements 562 We thank Daniel Brinkman (Yale Peabody Museum) for his assistance with the holotype 563 cervical vertebrae of Barosaurus, and Carl Mehling (American Museum of Natural 564 History) for out-of-hours access to the Hall of Saurischian Dinosaurs which houses the 565 vertebrae of the AMNH specimen. Jerry Harris (Dixie State University) provided a high- 566 resolution image of a Suuwassea vertebra. Several people left substantial and helpful 567 comments on the preprint of this paper: we thank Emanuel Tschopp (Universidade 568 Nova de Lisboa), Mark Robinson, Andrew Farke (Raymond M. Alf Museum), John 569 Foster (Museum of Moab) and Mickey Mortimer. 570
17 571 References 572 Barrett, Paul M. and Paul Upchurch Feeding mechanisms of Diplodocus. Gaia : Bartram, Alan, B. Booth, M. Chinery, E. N. K. Clarkson, B. Cox, D. Edwards, C. 575 Maynard and W. D. I. Rolfe The Prehistoric World. Galley Press (London). 576 Button, David, Emily Rayfield, and Paul Barrett Biomechanical evidence of niche 577 partitioning between sympatric sauropod dinosaurs. pp in Stig Walsh, Nick 578 Fraser, Stephen Brusatte, Jeff Liston and Vicen Carrió (eds.), Programme and 579 Abstracts, 61st Symposium on Vertebrae Palaeontology and Comparative Anatomy, 580 Edinburgh, UK, 27th 30th August pages. 581 Fiorillo, Anthony R Dental microwear patterns of the sauropod dinosaurs 582 Camarasaurus and Diplodocus: Evidence for resource partitioning in the Late 583 Jurassic of North America: Historical Biology 13: Foster, John R Comment #73 on Taylor and Wedel preprint, The neck of 585 Barosaurus was not only longer but also wider than those of Diplodocus and other 586 diplodocines (PeerJ PrePrints 1:e67v1) Gilmore, C. W Osteology of Apatosaurus with special reference to specimens in 588 the Carnegie Museum. Memoirs of the Carnegie Museum 11: Harris, Jerald D. 2006a. The axial skeleton of the dinosaur Suuwassea emilieae 590 (Sauropoda: Flagellicaudata) from the Upper Jurassic Morrison Formation of 591 Montana, USA. Palaeontology 49(5): Harris, Jerald D. 2006b. The significance of Suuwassea emiliae (Dinosauria: 593 Sauropoda) for flagellicaudatan intrarelationships and evolution. Journal of 594 Systematic Palaeontology 4: Hatcher, Jonathan B Diplodocus (Marsh): its osteology, taxonomy and probable 596 habits, with a restoration of the skeleton. Memoirs of the Carnegie Museum 1: and plates I XIII. 598 Hone, David W. E., Andrew A Farke and Mathew J. Wedel. Ontogeny and the fossil 599 record: what, if anything, is an adult dinosaur? Biology Letters 12(2): doi: /rsbl Janensch, Werner Die Wirbelsaule von Brachiosaurus brancai. 602 Palaeontographica (Suppl. 7) 3: Lambert, David DK Guide: Dinosaurs. Dorling Kindersley (London). 64 pages. 604 Lindsay, William Barosaurus: on the trail of the gigantic plant-eating dinosaur. 605 Dorling Kindersley (London). 32 pages. 606 Lull, Richard. S The sauropod dinosaur Barosaurus Marsh. Memoirs of the 607 Connecticut Academy of Arts and Sciences 6:1 42 and plates I VII.
18 608 Mannion, Philip D., Paul Upchurch, Octávio Mateus, Rosie N. Barnes & Marc E. H. 609 Jones New information on the anatomy and systematic position of 610 Dinheirosaurus lourinhanensis (Sauropoda: Diplodocoidea) from the Late Jurassic of 611 Portugal, with a review of European diplodocoids. Journal of Systematic 612 Palaeontology 10(3): doi: / Marsh, Othniel C Description of new dinosaurian reptiles. American Journal of 614 Science, 3rd series 39(229): Martin, John Mobility and feeding of Cetiosaurus (Saurischia, Sauropoda) why 616 the long neck? pp in P. J. Currie and E. H. Koster (eds), Fourth Symposium 617 on Mesozoic Terrestrial Ecosystems, Short Papers. Boxtree Books, Drumheller 618 (Alberta). 239 pages. 619 McIntosh, John S Sauropoda. pp in: D. B. Weishampel, P. Dodson and 620 H. Osmólska (eds.), The Dinosauria. University of California Press (Berkeley and Los 621 Angeles). 733 pages. 622 McIntosh, John S The Genus Barosaurus Marsh (Sauropoda, Diplodocidae). pp in Virginia Tidwell and Ken Carpenter (eds.), Thunder Lizards: the 624 Sauropodomorph Dinosaurs. Indiana University Press (Bloomington). 495 pages. 625 Novas, Fernando E., Leonardo Salgado, Jorge Calvo, and Federico Agnolin Giant titanosaur (Dinosauria, Sauropoda) from the Late Cretaceous of Patagonia. 627 Revisto del Museo Argentino de Ciencias Naturales, new series 7(1): O Gorman Eoin J., and David W. E. Hone Body size distribution of the dinosaurs. 629 PLOS ONE 7(12):e doi: /journal.pone Paul, Gregory S Terramegathermy and Cope's Rule in the land of titans. Modern 631 Geology 23: Paul, Gregory S The Scientific American Book of Dinosaurs. St. Martin's Press 633 (New York). 432 pages. 634 Ruxton, Graeme D. and David M. Wilkinson The energetics of low browsing in 635 sauropods. Biology Letters 7(5): doi: /rsbl Sander, P. Martin, Andreas Christian, Marcus Clauss, Regina Fechner, Carole T. Gee, 637 Eva-Maria Griebeler, Hanns-Christian Gunga, Jürgen Hummel, Heinrich Mallison, 638 Steven F. Perry, Holger Preuschoft, Oliver W. M. Rauhut, Kristian Remes, Thomas 639 Tütken, Oliver Wings and Ulrich Witzel Biology of the sauropod dinosaurs: the 640 evolution of gigantism. Biological Reviews 86: doi: /j X x 642 Senter, Phil Necks for sex: sexual selection as an explanation for sauropod 643 dinosaur neck elongation. Journal of Zoology 271(1): doi: /j x 645 Stevens, Kent A., and J. Michael Parrish Neck Posture and Feeding Habits of 646 Two Jurassic Sauropod Dinosaurs. Science 284:
19 647 Stevens, Kent A., and Michael J. Parrish. 2005a. Digital reconstructions of sauropod 648 dinosaurs and implications for feeding. pp in Jeffrey A. Wilson and Kristina 649 Curry Rogers (eds.), The Sauropods: Evolution and Paleobiology. University of 650 California Press (Berkeley). 363 pages. 651 Stevens, Kent A., and Michael J. Parrish. 2005b. Neck posture, dentition, and feeding 652 strategies in Jurassic sauropod dinosaurs. pp in Virginia Tidwell and Ken 653 Carpenter (eds.), Thunder Lizards: the Sauropodomorph Dinosaurs. Indiana 654 University Press (Bloomington, Indiana). 495 pages. 655 Taylor, Michael P Dinosaur diversity analysed by clade, age, place and year of 656 description. pp in Paul M. Barrett and Susan E. Evans (eds.), Ninth 657 international symposium on Mesozoic terrestrial ecosystems and biota, Manchester, 658 UK. Cambridge Publications. Natural History Museum, London, UK. 187 pp. 659 Taylor, Michael P. and Darren Naish The phylogenetic taxonomy of 660 Diplodocoidea (Dinosauria: Sauropoda). PaleoBios 25(2): Taylor, Michael P., and Mathew J. Wedel Why sauropods had long necks; and 662 why giraffes have short necks. PeerJ 1:e36. doi: /peerj Taylor, Michael P., Mathew J. Wedel and Darren Naish Head and neck posture 664 in sauropod dinosaurs inferred from extant animals. Acta Palaeontologica Polonica (2): Taylor, Michael P., David W. E. Hone, Mathew J. Wedel and Darren Naish. 2011a. The 667 long necks of sauropods did not evolve primarily through sexual selection. Journal of 668 Zoology 285: doi: /j x 669 Taylor, Michael P., Mathew J. Wedel and Richard L. Cifelli. 2011b. A new sauropod 670 dinosaur from the Lower Cretaceous Cedar Mountain Formation, Utah, USA. Acta 671 Palaeontologica Polonica 56(1): doi: /app Toon, A, and S. B. Toon Okapis and giraffes in: M. Hutchins, D. 673 Kleiman, V. Geist and M. McDade (eds.), Grzimek s Animal Life Encyclopedia, Vol : Mammals IV (second edition). Gale Group (Farmington Hills, Michigan). 675 Tschopp, Emanuel, and Octávio Mateus The skull and neck of a new 676 flagellicaudatan sauropod from the Morrison Formation and its implication for the 677 evolution and ontogeny of diplodocid dinosaurs. Journal of Systematic 678 Palaeontology, online ahead of print. doi: / Upchurch, Paul The phylogenetic relationships of sauropod dinosaurs. Zoological 680 Journal of the Linnean Society 124: Upchurch, Paul, Paul M. Barrett and Peter Dodson Sauropoda. pp in 682 D. B. Weishampel, P. Dodson and H. Osmólska (eds.), The Dinosauria, 2nd edition. 683 University of California Press (Berkeley and Los Angeles). 861 pages. 684 Upchurch, Paul, Yukimitsu Tomida, and Paul M. Barrett A new specimen of 685 Apatosaurus ajax (Sauropoda: Diplodocidae) from the Morrison Formation (Upper 686 Jurassic) of Wyoming, USA. National Science Museum Monographs No. 26. Tokyo pages.
20 688 van der Leeuw, Angelique H. J., Ron G. Bout, and Gart A. Zweers Evolutionary 689 morphology of the neck system in ratites, fowl, and waterfowl. Netherlands Journal of 690 Zoology 51(2): Wedel, Mathew J Postcranial skeletal pneumaticity in sauropods and its 692 implications for mass estimates. pp in Wilson, J. A., and Curry-Rogers, K. 693 (eds.), The Sauropods: Evolution and Paleobiology. University of California Press, 694 Berkeley. 695 Wedel, Mathew J Postcranial pneumaticity in dinosaurs and the origin of the 696 avian lung. Ph.D dissertation, Integrative Biology, University of California, Berkeley, 697 CA. Advisors: Kevin Padian and Bill Clemens. 304 pages. 698 Wedel, Mathew J., and R. Kent Sanders Osteological correlates of cervical 699 musculature in Aves and Sauropoda (Dinosauria: Saurischia), with comments on the 700 cervical ribs of Apatosaurus. PaleoBios 22(3): Wedel, Mathew J., and Michael P. Taylor Neural spine bifurcation in sauropod 702 dinosaurs of the Morrison Formation: ontogenetic and phylogenetic implications. 703 PalArch's Journal of Vertebrate Palaeontology 10(1): Wedel, Mathew J., Richard L. Cifelli and R. Kent Sanders. 2000a. Sauroposeidon 705 proteles, a new sauropod from the Early Cretaceous of Oklahoma. Journal of 706 Vertebrate Paleontology 20(1): Wedel, Mathew J., Richard L. Cifelli and R. Kent Sanders. 2000b. Osteology, 708 paleobiology, and relationships of the sauropod dinosaur Sauroposeidon. Acta 709 Palaeontologica Polonica 45(4): Whitlock, John A A phylogenetic analysis of Diplodocoidea (Saurischia: 711 Sauropoda). Zoological Journal of the Linnean Society 161(4): doi: /j x 713 Wilson, Jeffrey A Sauropod dinosaur phylogeny: critique and cladistic analysis. 714 Zoological Journal of the Linnean Society 136: Woodruff, D. Carey, and Denver W. Fowler Ontogenetic influence on neural 716 spine bifurcation in Diplodocoidea (Dinosauria: Sauropoda): a critical phylogenetic 717 character. Journal of Morphology 273(7): doi: /jmor Zweers Gart A., J. C. Vanden Berge, and R. Koppendraier Avian cranio-cervical 719 systems. Part I: Anatomy of the cervical column in the chicken (Gallus gallus L.). Acta 720 Morphologica Neerlando-Scandinavica 25:
21 722 Figure captions Figure 1. Mounted cast skeleton of Barosaurus referred specimen AMNH 6341, in the entrance hall of the American Museum of Natural History. Homo sapiens (MPT) for scale. Photograph by MJW. Figure 2. Posterior cervical vertebrae (C13) of diplodocine sauropods in lateral view, showing proportional differences. Top left, Kaatedocus, from Tschopp and Mateus (2012: unnumbered supplementary figure tjsp_a_746589_sup_ tif); top right, Barosaurus referred specimen AMNH 6341, left lateral, reversed (photo by MJW); bottom left, Diplodocus, from Hatcher (1901: plate III); bottom right, Diplodocus elongated by 33%. Kaatedocus and Barosaurus scaled to the same centrum length as original Diplodocus and elongated Diplodocus respectively. In lateral view, the widely assumed similarity between the cervicals of Barosaurus and elongated Diplodocus is largely borne out: the principal differences in Barosaurus are the less prominent and more posteriorly positioned neural spine (ns), the more prominent ventrolateral flanges (vlf), and the reduced pneumatic fossa (pf) in the centrum. Scale bars = 100 mm. Figure 3. Barosaurus lentus holotype YPM 429, vertebra R, C?15. Top row, left to right: posterior, dorsal and anterior views; middle row: right lateral view; bottom row: ventral view, from Lull (1919: plate II). Note the apparently very low, undivided neural spine at the intersection of the PRSLs and POSLs, forward-shifted neural arch, broad prezygapophyses, broad, wing-like prezygadiapophyseal laminae, and great width across the diapophyses and across the parapophyses. Abbreviations: dia, diapophysis; para, parapophysis; prz, prezygapophysis; prdl, prezygadiapophyseal lamina; spol, spinopostzygapophyseal lamina; sprl, spinoprezygapophyseal lamina. Scale bar = 500 mm. Figure 4. Diplodocid posterior vertebrae in dorsal view, scaled to equal total length, with neural spines highlighted. Left column, from top to bottom: Apatosaurus ajax Tokyo specimen NSMT- PV 20375, C12 (From Upchurch et al. 2005: plate I, part I), centrum length 380 mm; Barosaurus lentus holotype YPM 429, vertebra R (C?15), total length 960 mm. Right column, from top to bottom: Apatosaurus ajax holotype YPM 1860, C?11, centrum length unknown but probably about 500 mm; Suuwassea emilieae holotype ANS 21122, C7 (photograph supplied by Jerry Harris), centrum length 284 mm; Kaatedocus siberi holotype SMA 004, C13 (from unnumbered supplementary figure in Tschopp and Mateus 2012), total length 309 mm. Diplodocus is not pictured due to the lack of published illustrations. The vertebrae of Apatosaurus and Barosaurus are proportionally much wider than those of Suuwassea and Kaatedocus, and the bifurcation of the neural spine is far wider in both Apatosaurus specimens than in Suuwassea or Kaatedocus. No bifurcation is apparent in Barosaurus, which appears to have a low unsplit spine at the summit of four converging laminae, but this is a preservational artefact: see text. Scale bars = 100 mm. Figure 5. Giraffatitan brancai lectotype MB.R.2180, fifth cervical vertebra. Top row: left lateral view. Second row: dorsal view, with anterior to the right. Third row (from left to right): anterior, right lateral and posterior views. Bottom row: ventral view, with anterior to the right. Scale bar = 500 mm. Figure 6. Barosaurus lentus holotype YPM 429, Vertebra Q (C?13). Top row: left ventrolateral view. Middle row, from left to right: anterior view, with ventral to the right; ventral view; posterior view, with ventral to the left. Bottom row: right lateral view, inverted. Inset shows diapophyseal facet on right side of vertebra, indicating that the cervical ribs were unfused in this individual despite its great size. Note the broad, flat prezygapophyseal facet visible in anterior view. Scale
22 bar = 500 mm. Figure 7. Barosaurus lentus holotype YPM 429, Vertebra S (C?12). Left column from top to bottom: dorsal, right lateral and ventral views; right column: anterior view. Inset shows displaced fragment of broken prezygapophysis. Note the narrow span across the parapophyses in ventral view. Scale bar = 500 mm. Figure 8. Barosaurus lentus holotype YPM 429, cervical vertebrae in ventral view. From top to bottom: vertebra R (from Lull 1919: plate II), vertebra Q, vertebra S. Probably from more posterior to more anterior. Scale bar = 500 mm. Figure 9. Partial restoration of the Barosaurus lentus holotype YPM 429, cervical vertebra R, approximating its undamaged state by allowing for dorsoventral crushing, shearing and loss of some extremities. Anterior and posterior views scaled to 125% of uncorrected height and 80% of uncorrected width. Dorsal view scaled to 80% of uncorrected width; condyle moved forward and cotyle scaled to 50% of uncorrected width to allow for shearing. Lateral view scaled to 125% of uncorrected height, and sheared backwards 15 degrees. Lateral processes sheered upwards in anterior and posterior views. Metapophyses and postzygapophyses drawn in multiple views based on vertebrae Q and S, and C14 16 of AMNH Scale bar = 500 mm. Figure 10. Barosaurus AMNH 6341, cervical vertebrae in dorsal view, to scale. Left column, from top to bottom: C9 C12. Right column, from top to bottom: C14 C16. Extensive image manipulation was necessary to bring out the information in these photographs, due to to poor photography conditions. C16 is sheered to the right, so the aspect is slightly left dorsolateral rather than true dorsal. C8 is on display in the gallery with these vertebrae, but the structure of the display makes it impossible to photograph in dorsal view. C13 is on a shelf in collections, apart from the other cervicals, and we were not able to photograph it in dorsal view. Scale bar = 500 mm. Figure 11. Barosaurus AMNH 6341, cervical vertebrae C8 C16 in dorsal (where available) and lateral views, to scale. Lateral views except C13 from McIntosh (2005: fig. 2.1). Scale bar = 500 mm. Figure 12. Similarities between Barosaurus lentus holotype YPM 429, cervical vertebra R (C?15, left) and referred specimen AMNH 6341, C15 (right), scaled to same total length. Green brackets show width of prezygapophyseal rami, omitting apparent reconstruction on left anterolateral corner of YPM 429. Red outlines indicate margins of diapophyseal wings. Blue outlines show posterior fillets of diapophyseal wings. Orange X on AMNH 6341 indicates base of metapophyses, extended from prezygadiapophyseal and postzygadiapophyseal laminae and forming a diagonal cross similar to that of vertebra R. Prezygapophyseal facets of AMNH 6341 highlighted in yellow: the right facet is fairly clear in the photograph (see Figure 10); the exact margin of the left facet is less certain. Zygapophyseal facets cannot be directly recognised in vertebra R due to poor preservation and overzealous reconstruction. Scale bars = 500 mm. Figure 13. Attachments of the lateral flexor muscles of the neck in Kaatedocus and Barosaurus. On the left, C11 of Kaatedocus siberi holotype SMA 0004 (traced from Tschopp and Mateus 2012: fig. 10C2) in dorsal (top) and right lateral (bottom) views, with simplified versions of the lateral flexor muscles included, based on those of birds (see Wedel and Sanders 2002, and Taylor and Wedel 2013). The M. longus colli dorsalis and M. cervicalis ascendens insert together on the epipophysis (= torus dorsalis of birds), and the M. flexor colli lateralis and M. longus colli ventralis (ventral and medial, not shown) insert together on the cervical rib. The preepipophysis (sensu Tschopp and Mateus 2012) and the head of the cervical rib may have served as expanded attachments for M. cervicalis ascendens and M. flexor colli lateralis, respectively. The actual muscles were probably much more complex than those drawn here,
23 with numerous slips connecting multiple vertebrae: for a similar condition in birds, see Zweers et al. (1987) and van der Leeuw et al. (2001: fig. 2). On the right, C15 of Barosaurus AMNH 6341, scaled to the same total length as C11 of Kaatedocus. Actual total lengths for the two vertebrae are 840 mm for C15 of Barosaurus (McIntosh 2005: table 2.1) and 324 mm for C11 of Kaatedocus (Tschopp and Mateus 2012: table 1). In Barosaurus, the ansae costotransversariae or cervical rib loops are taller, wider and more posteriorly located than in Kaatedocus, providing a larger attachment area for the lateral flexor muscles (blue arcs) and lending them greater mechanical advantage (red lines). In this respect, Barosaurus is more similar to Apatosaurus than to the narrow-necked Diplodocus, although the cervical ribs of Barosaurus are much less robust than those of Apatosaurus.
24 Figure 1 Mounted cast skeleton of Barosaurus referred specimen AMNH 634. Mounted cast skeleton of Barosaurus referred specimen AMNH 6341, in the entrance hall of the American Museum of Natural History. Homo sapiens (MPT) for scale. Photograph by MJW.

25
26 Figure 2 Posterior cervical vertebrae (C13) of diplodocine sauropods in lateral view, showing proportional differences. Posterior cervical vertebrae (C13) of diplodocine sauropods in lateral view, showing proportional differences. Top left, Kaatedocus, from Tschopp and Mateus (2012: unnumbered supplementary figure tjsp_a_746589_sup_ tif); top right, Barosaurus referred specimen AMNH 6341, left lateral, reversed (photo by MJW); bottom left, Diplodocus, from Hatcher (1901: plate III); bottom right, Diplodocus elongated by 33%. Kaatedocus and Barosaurus scaled to the same centrum length as original Diplodocus and elongated Diplodocus respectively. In lateral view, the widely assumed similarity between the cervicals of Barosaurus and elongated Diplodocus is largely borne out: the principal differences in Barosaurus are the less prominent and more posteriorly positioned neural spine (ns), the more prominent ventrolateral flanges (vlf), and the reduced pneumatic fossa (pf) in the centrum. Scale bars = 100 mm.
27
28 Figure 3 Barosaurus lentus holotype YPM 429, vertebra R, C?15. Barosaurus lentus holotype YPM 429, vertebra R, C?15. Top row, left to right: posterior, dorsal and anterior views; middle row: right lateral view; bottom row: ventral view, from Lull (1919: plate II). Note the apparently very low, undivided neural spine at the intersection of the PRSLs and POSLs, forward-shifted neural arch, broad prezygapophyses, broad, wing-like prezygadiapophyseal laminae, and great width across the diapophyses and across the parapophyses. Abbreviations: dia, diapophysis; para, parapophysis; prz, prezygapophysis; prdl, prezygadiapophyseal lamina; spol, spinopostzygapophyseal lamina; sprl, spinoprezygapophyseal lamina. Scale bar = 500 mm.

29
30 Figure 4 Diplodocid posterior vertebrae in dorsal view, scaled to equal total length, with neural spines highlighted. Diplodocid posterior vertebrae in dorsal view, scaled to equal total length, with neural spines highlighted. Left column, from top to bottom: Apatosaurus ajax Tokyo specimen NSMT-PV 20375, C12 (From Upchurch et al. 2005: plate I, part I), centrum length 380 mm; Barosaurus lentus holotype YPM 429, vertebra R (C?15), total length 960 mm. Right column, from top to bottom: Apatosaurus ajax holotype YPM 1860, C?11, centrum length unknown but probably about 500 mm; Suuwassea emilieae holotype ANS 21122, C7 (photograph supplied by Jerry Harris), centrum length 284 mm; Kaatedocus siberi holotype SMA 004, C13 (from unnumbered supplementary figure in Tschopp and Mateus 2012), total length 309 mm. Diplodocus is not pictured due to the lack of published illustrations. The vertebrae of Apatosaurus and Barosaurus are proportionally much wider than those of Suuwassea and Kaatedocus, and the bifurcation of the neural spine is far wider in both Apatosaurus specimens than in Suuwassea or Kaatedocus. No bifurcation is apparent in Barosaurus, which appears to have a low unsplit spine at the summit of four converging laminae, but this is a preservational artefact: see text. Scale bars = 100 mm.
31
32 Figure 5 Giraffatitan brancai lectotype MB.R.2180, fifth cervical vertebra. Giraffatitan brancai lectotype MB.R.2180, fifth cervical vertebra. Top row: left lateral view. Second row: dorsal view, with anterior to the right. Third row (from left to right): anterior, right lateral and posterior views. Bottom row: ventral view, with anterior to the right. Scale bar = 500 mm.
33

34 Figure 6 Barosaurus lentus holotype YPM 429, Vertebra Q (C?13). Barosaurus lentus holotype YPM 429, Vertebra Q (C?13). Top row: left ventrolateral view. Middle row, from left to right: anterior view, with ventral to the right; ventral view; posterior view, with ventral to the left. Bottom row: right lateral view, inverted. Inset shows diapophyseal facet on right side of vertebra, indicating that the cervical ribs were unfused in this individual despite its great size. Note the broad, flat prezygapophyseal facet visible in anterior view. Scale bar = 500 mm.
35 Figure 7 Barosaurus lentus holotype YPM 429, Vertebra S (C?12). Barosaurus lentus holotype YPM 429, Vertebra S (C?12). Left column from top to bottom: dorsal, right lateral and ventral views; right column: anterior view. Inset shows displaced fragment of broken prezygapophysis. Note the narrow span across the parapophyses in ventral view. Scale bar = 500 mm.
36 Figure 8 Barosaurus lentus holotype YPM 429, cervical vertebrae in ventral view. Barosaurus lentus holotype YPM 429, cervical vertebrae in ventral view. From top to bottom: vertebra R (from Lull 1919: plate II), vertebra Q, vertebra S. Probably from more posterior to more anterior. Scale bar = 500 mm.

37
38 Figure 9 Partial restoration of the Barosaurus lentus holotype YPM 429. Partial restoration of the Barosaurus lentus holotype YPM 429, cervical vertebra R, approximating its undamaged state by allowing for dorsoventral crushing, shearing and loss of some extremities. Anterior and posterior views scaled to 125% of uncorrected height and 80% of uncorrected width. Dorsal view scaled to 80% of uncorrected width; condyle moved forward and cotyle scaled to 50% of uncorrected width to allow for shearing. Lateral view scaled to 125% of uncorrected height, and sheared backwards 15 degrees. Lateral processes sheered upwards in anterior and posterior views. Metapophyses and postzygapophyses drawn in multiple views based on vertebrae Q and S, and C14 16 of AMNH Scale bar = 500 mm.
39 Figure 10 Barosaurus AMNH 6341, cervical vertebrae in dorsal view, to scale. Barosaurus AMNH 6341, cervical vertebrae in dorsal view, to scale. Left column, from top to bottom: C9 C12. Right column, from top to bottom: C14 C16. Extensive image m
40
 and lateral views, to scale.")
41 Figure 11 Barosaurus AMNH 6341, cervical vertebrae C8 C16 in dorsal (where available) and lateral views, to scale. Barosaurus AMNH 6341, cervical vertebrae C8 C16 in dorsal (where available) and lateral views, to scale. Lateral views except C13 from McIntosh (2005: fig. 2.1). Scale bar = 500 mm.
42 Figure 12 Similarities between Barosaurus lentus holotype YPM 429, cervical vertebra R (C?15) and referred specimen AMNH 6341, C15. Similarities between Barosaurus lentus holotype YPM 429, cervical vertebra R (C?15, left) and referred specimen AMNH 6341, C15 (right), scaled to same total length. Green brackets show width of prezygapophyseal rami, omitting apparent reconstruction on left anterolateral corner of YPM 429. Red outlines indicate margins of diapophyseal wings. Blue outlines show posterior fillets of diapophyseal wings. Orange X on AMNH 6341 indicates base of metapophyses, extended from prezygadiapophyseal and postzygadiapophyseal laminae and forming a diagonal cross similar to that of vertebra R. Prezygapophyseal facets of AMNH 6341 highlighted in yellow: the right facet is fairly clear in the photograph (see Figure 10); the exact margin of the left facet is less certain. Zygapophyseal facets cannot be directly recognised in vertebra R due to poor preservation and overzealous reconstruction. Scale bars = 500 mm.
43
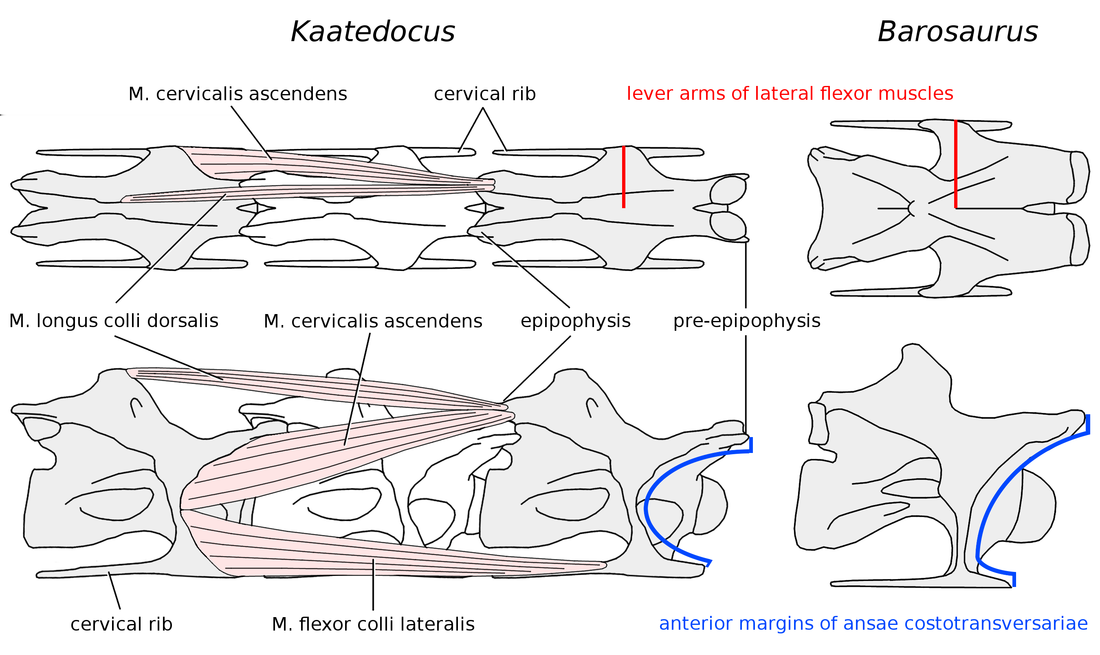
44 Figure 13 Attachments of the lateral flexor muscles of the neck in Kaatedocus and Barosaurus. Attachments of the lateral flexor muscles of the neck in Kaatedocus and Barosaurus. On the left, C11 of Kaatedocus siberi holotype SMA 0004 (traced from Tschopp and Mateus 2012: fig. 10C2) in dorsal (top) and right lateral (bottom) views, with simplified versions of the lateral flexor muscles included, based on those of birds (see Wedel and Sanders 2002, and Taylor and Wedel 2013). The M. longus colli dorsalis and M. cervicalis ascendens insert together on the epipophysis (= torus dorsalis of birds), and the M. flexor colli lateralis and M. longus colli ventralis (ventral and medial, not shown) insert together on the cervical rib. The preepipophysis (sensu Tschopp and Mateus 2012) and the head of the cervical rib may have served as expanded attachments for M. cervicalis ascendens and M. flexor colli lateralis, respectively. The actual muscles were probably much more complex than those drawn here, with numerous slips connecting multiple vertebrae: for a similar condition in birds, see Zweers et al. (1987) and van der Leeuw et al. (2001: fig. 2). On the right, C15 of Barosaurus AMNH 6341, scaled to the same total length as C11 of Kaatedocus. Actual total lengths for the two vertebrae are 840 mm for C15 of Barosaurus (McIntosh 2005: table 2.1) and 324 mm for C11 of Kaatedocus (Tschopp and Mateus 2012: table 1). In Barosaurus, the ansae costotransversariae or cervical rib loops are taller, wider and more posteriorly located than in Kaatedocus, providing a larger attachment area for the lateral flexor muscles (blue arcs) and lending them greater mechanical advantage (red lines). In this respect, Barosaurus is more similar to Apatosaurus than to the narrow-necked Diplodocus, although the cervical ribs of Barosaurus are much less robust than those of Apatosaurus.
45
Vertebral anatomy study guide. Human Structure Summer 2015. Prepared by Daniel Schmitt, Angel Zeininger, and Karyne Rabey.
 Vertebral anatomy study guide. Human Structure Summer 2015 Prepared by Daniel Schmitt, Angel Zeininger, and Karyne Rabey. 1. Plan of Action: In this guide you will learn to identify these structures: Cervical
Vertebral anatomy study guide. Human Structure Summer 2015 Prepared by Daniel Schmitt, Angel Zeininger, and Karyne Rabey. 1. Plan of Action: In this guide you will learn to identify these structures: Cervical
Almost all known sauropod necks are incomplete and distorted
 TAYLOR MOST SAUROPOD NECKS ARE INCOMPLETE (P1/19) Almost all known sauropod necks are incomplete and distorted Michael P. Taylor Department of Earth Sciences, University of Bristol, Bristol, England. dino@miketaylor.org.uk
TAYLOR MOST SAUROPOD NECKS ARE INCOMPLETE (P1/19) Almost all known sauropod necks are incomplete and distorted Michael P. Taylor Department of Earth Sciences, University of Bristol, Bristol, England. dino@miketaylor.org.uk
Lectures of Human Anatomy
 Lectures of Human Anatomy Vertebral Column-I By DR. ABDEL-MONEM AWAD HEGAZY M.B. with honor 1983, Dipl."Gynecology and Obstetrics "1989, Master "Anatomy and Embryology" 1994, M.D. "Anatomy and Embryology"
Lectures of Human Anatomy Vertebral Column-I By DR. ABDEL-MONEM AWAD HEGAZY M.B. with honor 1983, Dipl."Gynecology and Obstetrics "1989, Master "Anatomy and Embryology" 1994, M.D. "Anatomy and Embryology"
MORPHOLOGY OF A SPECIMEN OF SUPERSAURUS (DINOSAURIA, SAUROPODA) FROM THE MORRISON FORMATION OF WYOMING, AND A RE-EVALUATION OF DIPLODOCID PHYLOGENY 1
 FROM THE MORRISON FORMATION OF WYOMING, AND A RE-EVALUATION OF DIPLODOCID PHYLOGENY 1") Arquivos do Museu Nacional, Rio de Janeiro, v.65, n.4, p.527-544, out./dez.2007 ISSN 0365-4508 MORPHOLOGY OF A SPECIMEN OF SUPERSAURUS (DINOSAURIA, SAUROPODA) FROM THE MORRISON FORMATION OF WYOMING, AND
Arquivos do Museu Nacional, Rio de Janeiro, v.65, n.4, p.527-544, out./dez.2007 ISSN 0365-4508 MORPHOLOGY OF A SPECIMEN OF SUPERSAURUS (DINOSAURIA, SAUROPODA) FROM THE MORRISON FORMATION OF WYOMING, AND
THE DINOSAURS (CARNOSAURS, ALLOSAURIDS, SAUROPODS, CETIOSAURIDS) OF THE MIDDLE JURASSIC OF CERRO CÓNDOR (CHUBUT, ARGENTINA)
 OF THE MIDDLE JURASSIC OF CERRO CÓNDOR (CHUBUT, ARGENTINA)") Annales de Paléontologie (Vert.-Invert.) 1986, vol. 72, no. 4, pp. 325-386. THE DINOSAURS (CARNOSAURS, ALLOSAURIDS, SAUROPODS, CETIOSAURIDS) OF THE MIDDLE JURASSIC OF CERRO CÓNDOR (CHUBUT, ARGENTINA) by
Annales de Paléontologie (Vert.-Invert.) 1986, vol. 72, no. 4, pp. 325-386. THE DINOSAURS (CARNOSAURS, ALLOSAURIDS, SAUROPODS, CETIOSAURIDS) OF THE MIDDLE JURASSIC OF CERRO CÓNDOR (CHUBUT, ARGENTINA) by
AN UNUSUAL NEW NEOSAUROPOD DINOSAUR FROM THE LOWER CRETACEOUS HASTINGS BEDS GROUP OF EAST SUSSEX, ENGLAND
 [Palaeontology, Vol. 50, Part 6, 2007, pp. 1547 1564] AN UNUSUAL NEW NEOSAUROPOD DINOSAUR FROM THE LOWER CRETACEOUS HASTINGS BEDS GROUP OF EAST SUSSEX, ENGLAND by MICHAEL P. TAYLOR and DARREN NAISH Palaeobiology
[Palaeontology, Vol. 50, Part 6, 2007, pp. 1547 1564] AN UNUSUAL NEW NEOSAUROPOD DINOSAUR FROM THE LOWER CRETACEOUS HASTINGS BEDS GROUP OF EAST SUSSEX, ENGLAND by MICHAEL P. TAYLOR and DARREN NAISH Palaeobiology
Human Anatomy & Physiology
 PowerPoint Lecture Slides prepared by Barbara Heard, Atlantic Cape Community College Ninth Edition Human Anatomy & Physiology C H A P T E R 7 The Skeleton: Part B Annie Leibovitz/Contact Press Images Vertebral
PowerPoint Lecture Slides prepared by Barbara Heard, Atlantic Cape Community College Ninth Edition Human Anatomy & Physiology C H A P T E R 7 The Skeleton: Part B Annie Leibovitz/Contact Press Images Vertebral
Erketu ellisoni, a Long-Necked Sauropod from Bor Guvé (Dornogov Aimag, Mongolia)
") PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3508, 16 pp., 12 figures, 3 tables March 16, 2006 Erketu ellisoni, a Long-Necked Sauropod
PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3508, 16 pp., 12 figures, 3 tables March 16, 2006 Erketu ellisoni, a Long-Necked Sauropod
Clarification of Terms
 Shoulder Girdle Clarification of Terms Shoulder girdle = scapula and clavicle Shoulder joint (glenohumeral joint) = scapula and humerus What is the purpose (or function) of the shoulder and entire upper
Shoulder Girdle Clarification of Terms Shoulder girdle = scapula and clavicle Shoulder joint (glenohumeral joint) = scapula and humerus What is the purpose (or function) of the shoulder and entire upper
Anatomy and Pathomechanics of the Sacrum and Pelvis. Charles R. Thompson Head Athletic Trainer Princeton University
 Anatomy and Pathomechanics of the Sacrum and Pelvis Charles R. Thompson Head Athletic Trainer Princeton University Simplify Everything There are actually only three bones: Two innominates, one sacrum.
Anatomy and Pathomechanics of the Sacrum and Pelvis Charles R. Thompson Head Athletic Trainer Princeton University Simplify Everything There are actually only three bones: Two innominates, one sacrum.
DEVELOPMENT AND GROWTH OF THE MANDIBLE
 2012-2013 ORAL BIOLOGY DEVELOPMENT AND GROWTH OF THE MANDIBLE Ass. Prof. Dr. Heba M. Elsabaa Development and Growth of the Mandible DEVELOPMENT OF THE MANDIBLE The Mandible Is the largest and strongest
2012-2013 ORAL BIOLOGY DEVELOPMENT AND GROWTH OF THE MANDIBLE Ass. Prof. Dr. Heba M. Elsabaa Development and Growth of the Mandible DEVELOPMENT OF THE MANDIBLE The Mandible Is the largest and strongest
IDENTIFICATION OF ADULT CORIXIDS 25
 IDENTIFICATION OF ADULT CORIXIDS 25 THE IDENTIFICATION OF BRITISH ADULT SPECIMENS OF SIGARA LATERALIS (LEACH), SIGARA CONCINNA (FIEBER), CALLICORIXA PRAEUSTA (FIEBER) AND CALLICORIXA WOLLASTONI (DOUGLAS
IDENTIFICATION OF ADULT CORIXIDS 25 THE IDENTIFICATION OF BRITISH ADULT SPECIMENS OF SIGARA LATERALIS (LEACH), SIGARA CONCINNA (FIEBER), CALLICORIXA PRAEUSTA (FIEBER) AND CALLICORIXA WOLLASTONI (DOUGLAS
DESCRIPTION OF A TITANOSAURID CAUDAL SERIES FROM THE BAURU GROUP, LATE CRETACEOUS OF BRAZIL 1
 Arquivos do Museu Nacional, Rio de Janeiro, v.63, n.3, p.529-564, jul./set.2005 ISSN 0365-4508 DESCRIPTION OF A TITANOSAURID CAUDAL SERIES FROM THE BAURU GROUP, LATE CRETACEOUS OF BRAZIL 1 (With 27 figures)
Arquivos do Museu Nacional, Rio de Janeiro, v.63, n.3, p.529-564, jul./set.2005 ISSN 0365-4508 DESCRIPTION OF A TITANOSAURID CAUDAL SERIES FROM THE BAURU GROUP, LATE CRETACEOUS OF BRAZIL 1 (With 27 figures)
Lumbar Spine Anatomy. eorthopod.com 228 West Main St., Suite D Missoula, MT 59802-4345 Phone: 406-721-3072 Fax: 406-721-2619 info@eorthopod.
 A Patient s Guide to Lumbar Spine Anatomy 228 West Main St., Suite D Missoula, MT 59802-4345 Phone: 406-721-3072 Fax: 406-721-2619 info@eorthopod.com DISCLAIMER: The information in this booklet is compiled
A Patient s Guide to Lumbar Spine Anatomy 228 West Main St., Suite D Missoula, MT 59802-4345 Phone: 406-721-3072 Fax: 406-721-2619 info@eorthopod.com DISCLAIMER: The information in this booklet is compiled
NECK POSTURE, DENTITION, AND FEEDING STRATEGIES IN JURASSIC SAUROPOD DINOSAURS. KENT A. STEVENS and J. MICHAEL PARRISH
 NECK POSTURE, DENTITION, AND FEEDING STRATEGIES IN JURASSIC SAUROPOD DINOSAURS KENT A. STEVENS and J. MICHAEL PARRISH Department of Computer and Information Science University of Oregon, Eugene, OR 97403;
NECK POSTURE, DENTITION, AND FEEDING STRATEGIES IN JURASSIC SAUROPOD DINOSAURS KENT A. STEVENS and J. MICHAEL PARRISH Department of Computer and Information Science University of Oregon, Eugene, OR 97403;
THE BENJAMIN INSTITUTE PRESENTS. Excerpt from Listen To Your Pain. Assessment & Treatment of. Low Back Pain. Ben E. Benjamin, Ph.D.
 THE BENJAMIN INSTITUTE PRESENTS Excerpt from Listen To Your Pain Assessment & Treatment of Low Back Pain A B E N J A M I N I N S T I T U T E E B O O K Ben E. Benjamin, Ph.D. 2 THERAPIST/CLIENT MANUAL The
THE BENJAMIN INSTITUTE PRESENTS Excerpt from Listen To Your Pain Assessment & Treatment of Low Back Pain A B E N J A M I N I N S T I T U T E E B O O K Ben E. Benjamin, Ph.D. 2 THERAPIST/CLIENT MANUAL The
Disc Replacement or Fusion: which is best for me?
 Disc Replacement or Fusion: which is best for me? After anterior discectomy in either the cervical or lumbar region there is a need to replace the removed disc with something and the choice lies between
Disc Replacement or Fusion: which is best for me? After anterior discectomy in either the cervical or lumbar region there is a need to replace the removed disc with something and the choice lies between
Table S1. List of Institutional Abbreviations. AMNH, American Museum of Natural History, New York, New York; ANSP, Academy of National Science of
 Table S1. List of Institutional Abbreviations. AMNH, American Museum of Natural History, New York, New York; ANSP, Academy of National Science of Philadelphia, Philadelphia, Pennsylvania; BHI, Black Hills
Table S1. List of Institutional Abbreviations. AMNH, American Museum of Natural History, New York, New York; ANSP, Academy of National Science of Philadelphia, Philadelphia, Pennsylvania; BHI, Black Hills
Lab 5 Overview of the Skeleton: Classification and Structure of Bones and Cartilages Exercise 9 The Axial Skeleton Exercise 10
 Lab 5 Overview of the Skeleton: Classification and Structure of Bones and Cartilages Exercise 9 The Axial Skeleton Exercise 10 Overview of the Skeleton Locate the important cartilages in the human skeleton
Lab 5 Overview of the Skeleton: Classification and Structure of Bones and Cartilages Exercise 9 The Axial Skeleton Exercise 10 Overview of the Skeleton Locate the important cartilages in the human skeleton
Full Crown Module: Learner Level 1
 Full Crown Module Restoration / Tooth # Full Gold Crown (FGC) / 30 Extensions: Porcelain Fused to Metal (PFM) / 12 All Ceramic / 8 Learner Level 1 Mastery of Tooth Preparation Estimated Set Up Time: 30
Full Crown Module Restoration / Tooth # Full Gold Crown (FGC) / 30 Extensions: Porcelain Fused to Metal (PFM) / 12 All Ceramic / 8 Learner Level 1 Mastery of Tooth Preparation Estimated Set Up Time: 30
Section 2 Scientific Methods in Earth Science
 Section 2 Scientific Methods in Earth Science Key Concept Scientists conduct careful investigations by following standard methods that allow them to collect data and communicate results. What You Will
Section 2 Scientific Methods in Earth Science Key Concept Scientists conduct careful investigations by following standard methods that allow them to collect data and communicate results. What You Will
Session 7 Bivariate Data and Analysis
 Session 7 Bivariate Data and Analysis Key Terms for This Session Previously Introduced mean standard deviation New in This Session association bivariate analysis contingency table co-variation least squares
Session 7 Bivariate Data and Analysis Key Terms for This Session Previously Introduced mean standard deviation New in This Session association bivariate analysis contingency table co-variation least squares
Implants in your Laboratory: Abutment Design
 1/2 point CDT documented scientific credit. See Page 41. Implants in your Laboratory: Abutment Design By Leon Hermanides, CDT A patient s anatomical limitations have the greatest predictive value for successful
1/2 point CDT documented scientific credit. See Page 41. Implants in your Laboratory: Abutment Design By Leon Hermanides, CDT A patient s anatomical limitations have the greatest predictive value for successful
Thoracic Spine Anatomy
 A Patient s Guide to Thoracic Spine Anatomy 228 West Main, Suite C Missoula, MT 59802 Phone: info@spineuniversity.com DISCLAIMER: The information in this booklet is compiled from a variety of sources.
A Patient s Guide to Thoracic Spine Anatomy 228 West Main, Suite C Missoula, MT 59802 Phone: info@spineuniversity.com DISCLAIMER: The information in this booklet is compiled from a variety of sources.
Jack Russell /Parson Russell Terrier Spanning Guide
 Jack Russell /Parson Russell Terrier Spanning Guide One of the most important attributes of a Jack Russell Terrier is his spannable, flexible chest. Equally important is the shape of his chest. It is this
Jack Russell /Parson Russell Terrier Spanning Guide One of the most important attributes of a Jack Russell Terrier is his spannable, flexible chest. Equally important is the shape of his chest. It is this
Dental Updates. Excerpted Article e-mail: re777@comcast.net. Why Implant Screws Loosen Part 1. Richard Erickson, MS, DDS
 ¼ ½ ¾ µ mw/cm 2 Volume 17; 2007 Dental Updates "CUTTING EDGE INFORMATION FOR THE DENTAL PROFESSIONAL " 200 SEMINARS AND 30 JOURNALS REVIEWED YEARLY FOR THE LATEST, CUTTING EDGE INFORMATION Excerpted Article
¼ ½ ¾ µ mw/cm 2 Volume 17; 2007 Dental Updates "CUTTING EDGE INFORMATION FOR THE DENTAL PROFESSIONAL " 200 SEMINARS AND 30 JOURNALS REVIEWED YEARLY FOR THE LATEST, CUTTING EDGE INFORMATION Excerpted Article
JACK RUSSELL TERRIER
 05.12.2012 / EN FEDERATION CYNOLOGIQUE INTERNATIONALE (AISBL) SECRETARIAT GENERAL: 13, Place Albert 1 er B 6530 Thuin (Belgique) FCI-Standard N 345 JACK RUSSELL TERRIER SMOOTH COATED VARIETY BROKEN COATED
05.12.2012 / EN FEDERATION CYNOLOGIQUE INTERNATIONALE (AISBL) SECRETARIAT GENERAL: 13, Place Albert 1 er B 6530 Thuin (Belgique) FCI-Standard N 345 JACK RUSSELL TERRIER SMOOTH COATED VARIETY BROKEN COATED
Divisions of the Skeletal System
 OpenStax-CNX module: m46344 1 Divisions of the Skeletal System OpenStax College This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the end of this
OpenStax-CNX module: m46344 1 Divisions of the Skeletal System OpenStax College This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the end of this
Dynamics of dinosaurs
 MATH0011 Numbers and Patterns in Nature and Life Lecture 1 Dynamics of Dinosaurs http://147.8.101.93/math0011/ Questions: Dynamics of dinosaurs How heavy are dinosaurs? Would sand support a big dinosaur
MATH0011 Numbers and Patterns in Nature and Life Lecture 1 Dynamics of Dinosaurs http://147.8.101.93/math0011/ Questions: Dynamics of dinosaurs How heavy are dinosaurs? Would sand support a big dinosaur
Muscular System. Student Learning Objectives: Identify the major muscles of the body Identify the action of major muscles of the body
 Muscular System Student Learning Objectives: Identify the major muscles of the body Identify the action of major muscles of the body Structures to be identified: Muscle actions: Extension Flexion Abduction
Muscular System Student Learning Objectives: Identify the major muscles of the body Identify the action of major muscles of the body Structures to be identified: Muscle actions: Extension Flexion Abduction
Chapter 4 The Shoulder Girdle
 Chapter 4 The Shoulder Girdle Key Manubrium Clavicle Coracoidprocess Acromionprocess bony landmarks Glenoid fossa Bones Lateral Inferior Medial border angle McGraw-Hill Higher Education. All rights reserved.
Chapter 4 The Shoulder Girdle Key Manubrium Clavicle Coracoidprocess Acromionprocess bony landmarks Glenoid fossa Bones Lateral Inferior Medial border angle McGraw-Hill Higher Education. All rights reserved.
Anatomy and Terminology of the Spine. Bones of the Spine (Vertebrae)
") Anatomy and Terminology of the Spine The spine, also called the spinal column, vertebral column or backbone, consists of bones, intervertebral discs, ligaments, and joints. In addition, the spine serves
Anatomy and Terminology of the Spine The spine, also called the spinal column, vertebral column or backbone, consists of bones, intervertebral discs, ligaments, and joints. In addition, the spine serves
The Anatomy of Spinal Cord Injury (SCI)
") The Anatomy of Spinal Cord Injury (SCI) What is the Spinal Cord? The spinal cord is that part of your central nervous system that transmits messages between your brain and your body. The spinal cord has
The Anatomy of Spinal Cord Injury (SCI) What is the Spinal Cord? The spinal cord is that part of your central nervous system that transmits messages between your brain and your body. The spinal cord has
Laerdal' Human Anatomy Manual The Skeleton
 Human Anatomy Manual The Skeleton Laerdal Texas P.O. Box 38.226 EM. 116 Gatesville,Texas U.S.A.76528 U.S.A.1-800-433-5539 IntemationaI1-254-865-7221 24 Hour Fax 254-865-8011 ~ Laerdal' TABLE OF CONTENTS
Human Anatomy Manual The Skeleton Laerdal Texas P.O. Box 38.226 EM. 116 Gatesville,Texas U.S.A.76528 U.S.A.1-800-433-5539 IntemationaI1-254-865-7221 24 Hour Fax 254-865-8011 ~ Laerdal' TABLE OF CONTENTS
Unit 4: Skeletal System Test Review Test Review
 Name: Period: Unit 4: Skeletal System Test Review Test Review 1. List four functions of the skeletal system: a. b. c. d. 2. Define ossification and identify the roles of the osteoblasts, osteocytes, and
Name: Period: Unit 4: Skeletal System Test Review Test Review 1. List four functions of the skeletal system: a. b. c. d. 2. Define ossification and identify the roles of the osteoblasts, osteocytes, and
SPINAL FUSION. North American Spine Society Public Education Series
 SPINAL FUSION North American Spine Society Public Education Series WHAT IS SPINAL FUSION? The spine is made up of a series of bones called vertebrae ; between each vertebra are strong connective tissues
SPINAL FUSION North American Spine Society Public Education Series WHAT IS SPINAL FUSION? The spine is made up of a series of bones called vertebrae ; between each vertebra are strong connective tissues
Mini Dinosaurs. Grades K 1 2. Compiled by
 Mini Dinosaurs Grades K 1 2 Compiled by Leanne M. Burrow, Extension educator, 4 H/ Youth development, Purdue University Cooperative Extension Service Henry County Resources and References Exploring 4 H,
Mini Dinosaurs Grades K 1 2 Compiled by Leanne M. Burrow, Extension educator, 4 H/ Youth development, Purdue University Cooperative Extension Service Henry County Resources and References Exploring 4 H,
On Cervical Zygapophysial Joint Pain After Whiplash. Spine December 1, 2011; Volume 36, Number 25S, pp S194 S199
 On Cervical Zygapophysial Joint Pain After Whiplash 1 Spine December 1, 2011; Volume 36, Number 25S, pp S194 S199 Nikolai Bogduk, MD, PhD FROM ABSTRACT Objective To summarize the evidence that implicates
On Cervical Zygapophysial Joint Pain After Whiplash 1 Spine December 1, 2011; Volume 36, Number 25S, pp S194 S199 Nikolai Bogduk, MD, PhD FROM ABSTRACT Objective To summarize the evidence that implicates
OFFICE FOR HARMONIZATION IN THE INTERNAL MARKET (TRADE MARKS AND DESIGNS) DECISION of the First Board of Appeal of 8 July 2010
 DECISION of the First Board of Appeal of 8 July 2010") OFFICE FOR HARMONIZATION IN THE INTERNAL MARKET (TRADE MARKS AND DESIGNS) The Boards of Appeal DECISION of the First Board of Appeal of 8 July 2010 In Case R 366/2010-1 CONTESSA PREMIUM FOODS, INC. 222
OFFICE FOR HARMONIZATION IN THE INTERNAL MARKET (TRADE MARKS AND DESIGNS) The Boards of Appeal DECISION of the First Board of Appeal of 8 July 2010 In Case R 366/2010-1 CONTESSA PREMIUM FOODS, INC. 222
Anatomy and Physiology 121: Muscles of the Human Body
 Epicranius Anatomy and Physiology 121: Muscles of the Human Body Covers upper cranium Raises eyebrows, surprise, headaches Parts Frontalis Occipitalis Epicranial aponeurosis Orbicularis oculi Ring (sphincter)
Epicranius Anatomy and Physiology 121: Muscles of the Human Body Covers upper cranium Raises eyebrows, surprise, headaches Parts Frontalis Occipitalis Epicranial aponeurosis Orbicularis oculi Ring (sphincter)
Problems. Knowing. back of the
 Introduction Welcome to BodyZone Physiotherapy patient resource about Cervical Spine Problems. Knowing the main parts of your neck and how these parts work is important as you learn to care for your neck
Introduction Welcome to BodyZone Physiotherapy patient resource about Cervical Spine Problems. Knowing the main parts of your neck and how these parts work is important as you learn to care for your neck
The Avian Skeleton. Avian Flight. The Pelvic Girdle. Skeletal Strength. The Pelvic Girdle
 Flight is the central avian adaptation Birds can fly at great speeds, for extended distances and time periods, can soar for days, and hover, fly upside down, and backwards Adaptations for flight are integrated
Flight is the central avian adaptation Birds can fly at great speeds, for extended distances and time periods, can soar for days, and hover, fly upside down, and backwards Adaptations for flight are integrated
Analysis of academy school performance in GCSEs 2014
 Analysis of academy school performance in GCSEs 2014 Final report Report Analysis of academy school performance in GCSEs 2013 1 Analysis of Academy School Performance in GCSEs 2014 Jack Worth Published
Analysis of academy school performance in GCSEs 2014 Final report Report Analysis of academy school performance in GCSEs 2013 1 Analysis of Academy School Performance in GCSEs 2014 Jack Worth Published
FORENSIC ANTHROPOLOGY NOTES
 FORENSIC ANTHROPOLOGY NOTES Forensic Anthropology = the examinations of human skeletal remains for law enforcement agencies to determine the identity of unidentified bones. Questions asked: 1. Are the
FORENSIC ANTHROPOLOGY NOTES Forensic Anthropology = the examinations of human skeletal remains for law enforcement agencies to determine the identity of unidentified bones. Questions asked: 1. Are the
Stability of the spine modelled as an arch
 Loughborough University Institutional Repository Stability of the spine modelled as an arch This item was submitted to Loughborough University's Institutional Repository by the/an author. Citation: XIAO,
Loughborough University Institutional Repository Stability of the spine modelled as an arch This item was submitted to Loughborough University's Institutional Repository by the/an author. Citation: XIAO,
Observing Vertebrate Skeletons
 Name Class Date Chapter 33 Comparing Chordates Observing Vertebrate Skeletons Introduction One characteristic common to all vertebrates is the presence of a skeleton. The endoskeleton provides support,
Name Class Date Chapter 33 Comparing Chordates Observing Vertebrate Skeletons Introduction One characteristic common to all vertebrates is the presence of a skeleton. The endoskeleton provides support,
Evidence for evolution factsheet
 The theory of evolution by natural selection is supported by a great deal of evidence. Fossils Fossils are formed when organisms become buried in sediments, causing little decomposition of the organism.
The theory of evolution by natural selection is supported by a great deal of evidence. Fossils Fossils are formed when organisms become buried in sediments, causing little decomposition of the organism.
Copyright 2011 Casa Software Ltd. www.casaxps.com. Centre of Mass
 Centre of Mass A central theme in mathematical modelling is that of reducing complex problems to simpler, and hopefully, equivalent problems for which mathematical analysis is possible. The concept of
Centre of Mass A central theme in mathematical modelling is that of reducing complex problems to simpler, and hopefully, equivalent problems for which mathematical analysis is possible. The concept of
BIOL 4260 Human Evolu3onary Anatomy Lecture 5: Bone Development & Trunk Anatomy. Lecture 2: Fossil Record
 BIOL 4260 Human Evolu3onary Anatomy Lecture 5: Bone Development & Trunk Anatomy Lecture 2: Fossil Record Segmentation Cyclic genescreate segme ntation clock for somite production Final #s 4 occipital 8
BIOL 4260 Human Evolu3onary Anatomy Lecture 5: Bone Development & Trunk Anatomy Lecture 2: Fossil Record Segmentation Cyclic genescreate segme ntation clock for somite production Final #s 4 occipital 8
ANTERIOR CERVICAL DISCECTOMY AND FUSION. Basic Anatomical Landmarks: Anterior Cervical Spine
 Anterior In the human anatomy, referring to the front surface of the body or position of one structure relative to another Cervical Relating to the neck, in the spine relating to the first seven vertebrae
Anterior In the human anatomy, referring to the front surface of the body or position of one structure relative to another Cervical Relating to the neck, in the spine relating to the first seven vertebrae
INTERNATIONAL COMPARISONS OF PART-TIME WORK
 OECD Economic Studies No. 29, 1997/II INTERNATIONAL COMPARISONS OF PART-TIME WORK Georges Lemaitre, Pascal Marianna and Alois van Bastelaer TABLE OF CONTENTS Introduction... 140 International definitions
OECD Economic Studies No. 29, 1997/II INTERNATIONAL COMPARISONS OF PART-TIME WORK Georges Lemaitre, Pascal Marianna and Alois van Bastelaer TABLE OF CONTENTS Introduction... 140 International definitions
2. Incidence, prevalence and duration of breastfeeding
 2. Incidence, prevalence and duration of breastfeeding Key Findings Mothers in the UK are breastfeeding their babies for longer with one in three mothers still breastfeeding at six months in 2010 compared
2. Incidence, prevalence and duration of breastfeeding Key Findings Mothers in the UK are breastfeeding their babies for longer with one in three mothers still breastfeeding at six months in 2010 compared
Animal daoyin. Dr. Shen Hongxun. Exercises of the BUQI System
 Dr. Shen Hongxun Animal daoyin Exercises of the BUQI System Copyright 2005; Dr. Shen Hongxun, Gent All rights reserved. This tekst may not be reproduced, in whole or in part, by photocopy or print or any
Dr. Shen Hongxun Animal daoyin Exercises of the BUQI System Copyright 2005; Dr. Shen Hongxun, Gent All rights reserved. This tekst may not be reproduced, in whole or in part, by photocopy or print or any
giganotosaurus Michael P. Goecke
 giganotosaurus Michael P. Goecke Giganotosaurus A Buddy Book by Michael P. Goecke VISIT US AT www.abdopublishing.com Published by ABDO Publishing Company, 4940 Viking Drive, Edina, Minnesota 55435. Copyright
giganotosaurus Michael P. Goecke Giganotosaurus A Buddy Book by Michael P. Goecke VISIT US AT www.abdopublishing.com Published by ABDO Publishing Company, 4940 Viking Drive, Edina, Minnesota 55435. Copyright
Skeletal System. Axial Skeleton: Vertebral Column and Ribs
 Skeletal System Axial Skeleton: Vertebral Column and Ribs Functions Regions Cervical Thoracic Lumbar Sacral Primary & secondary curvatures There are three major functions of the vertebral column. First,
Skeletal System Axial Skeleton: Vertebral Column and Ribs Functions Regions Cervical Thoracic Lumbar Sacral Primary & secondary curvatures There are three major functions of the vertebral column. First,
Biomechanical Analysis of the Deadlift (aka Spinal Mechanics for Lifters) Tony Leyland
 Tony Leyland") Biomechanical Analysis of the Deadlift (aka Spinal Mechanics for Lifters) Tony Leyland Mechanical terminology The three directions in which forces are applied to human tissues are compression, tension,
Biomechanical Analysis of the Deadlift (aka Spinal Mechanics for Lifters) Tony Leyland Mechanical terminology The three directions in which forces are applied to human tissues are compression, tension,
3D Drawing. Single Point Perspective with Diminishing Spaces
 3D Drawing Single Point Perspective with Diminishing Spaces The following document helps describe the basic process for generating a 3D representation of a simple 2D plan. For this exercise we will be
3D Drawing Single Point Perspective with Diminishing Spaces The following document helps describe the basic process for generating a 3D representation of a simple 2D plan. For this exercise we will be
(J~ GREGORY S. PAUL 3109 N. Calvert St. Side Apt. Baltimore, MD 21218 ABSTRACT
 \ \\.,.., vol. 2 llo.3 HUNTERIA Jocit:ta& (J ISSN No. 0892-3701, University of Colorado Museum, Campus Box 315, Boulder, CO 80309-0315 THE BRACHIOSAUR GIANTS OF THE MORRISON AND TENDAGURU WITH A DESCRIPTION
\ \\.,.., vol. 2 llo.3 HUNTERIA Jocit:ta& (J ISSN No. 0892-3701, University of Colorado Museum, Campus Box 315, Boulder, CO 80309-0315 THE BRACHIOSAUR GIANTS OF THE MORRISON AND TENDAGURU WITH A DESCRIPTION
Feathered Dinosaurs and the Origin of Flight
 Feathered Dinosaurs and the Origin of Flight Exhibition Organized and Circulated by: The Dinosaur Museum, Utah The Fossil Administration Office, Liaoning, China Beipiao City Paleontological Research Center,
Feathered Dinosaurs and the Origin of Flight Exhibition Organized and Circulated by: The Dinosaur Museum, Utah The Fossil Administration Office, Liaoning, China Beipiao City Paleontological Research Center,
Biomechanics of Joints, Ligaments and Tendons.
 Hippocrates (460-377 B.C.) Biomechanics of Joints, s and Tendons. Course Text: Hamill & Knutzen (some in chapter 2 and 3, but ligament and tendon mechanics is not well covered in the text) Nordin & Frankel
Hippocrates (460-377 B.C.) Biomechanics of Joints, s and Tendons. Course Text: Hamill & Knutzen (some in chapter 2 and 3, but ligament and tendon mechanics is not well covered in the text) Nordin & Frankel
Anatomy PHL 212. By Dr Tajdar Husain Khan
 Anatomy PHL 212 By Dr Tajdar Husain Khan Overview of Anatomy Anatomy(from the Greek word anatome,"dissection") is a branch of natural science dealing with the structural organization of living things The
Anatomy PHL 212 By Dr Tajdar Husain Khan Overview of Anatomy Anatomy(from the Greek word anatome,"dissection") is a branch of natural science dealing with the structural organization of living things The
Gallina & Apesteguia: Upper Cretaceous rebbachisaurid from Northern Patagonia 153
 Gallina & Apesteguia: Upper Cretaceous rebbachisaurid from Northern Patagonia 153 Rev. Mus. Argentino Cienc. Nat., n.s. 7(2): 153-166, 2005 Buenos Aires, ISSN 1514-5158 Cathartesaura anaerobica gen. et
Gallina & Apesteguia: Upper Cretaceous rebbachisaurid from Northern Patagonia 153 Rev. Mus. Argentino Cienc. Nat., n.s. 7(2): 153-166, 2005 Buenos Aires, ISSN 1514-5158 Cathartesaura anaerobica gen. et
BASIC EXERCISES AND DRESSAGE MOVEMENTS
 Appendix USDF Teaching Manual Articles BASIC EXERCISES AND DRESSAGE MOVEMENTS Excerpted from Principles of Riding The Official Handbook of the German National Equestrian Federation, 1985 edition Reprinted
Appendix USDF Teaching Manual Articles BASIC EXERCISES AND DRESSAGE MOVEMENTS Excerpted from Principles of Riding The Official Handbook of the German National Equestrian Federation, 1985 edition Reprinted
Objectives AXIAL SKELETON. 1. Frontal Bone. 2. Parietal Bones. 3. Temporal Bones. CRANIAL BONES (8 total flat bones w/ 2 paired)
") Objectives AXIAL SKELETON SKULL 1. On a skull or diagram, identify and name the bones of the skull 2. Identify the structure and function of the bones of the skull 3. Describe how a fetal skull differs
Objectives AXIAL SKELETON SKULL 1. On a skull or diagram, identify and name the bones of the skull 2. Identify the structure and function of the bones of the skull 3. Describe how a fetal skull differs
Sample Size and Power in Clinical Trials
 Sample Size and Power in Clinical Trials Version 1.0 May 011 1. Power of a Test. Factors affecting Power 3. Required Sample Size RELATED ISSUES 1. Effect Size. Test Statistics 3. Variation 4. Significance
Sample Size and Power in Clinical Trials Version 1.0 May 011 1. Power of a Test. Factors affecting Power 3. Required Sample Size RELATED ISSUES 1. Effect Size. Test Statistics 3. Variation 4. Significance
Writing a degree project at Lund University student perspectives
 1 Writing a degree project at Lund University student perspectives Summary This report summarises the results of a survey that focused on the students experiences of writing a degree project at Lund University.
1 Writing a degree project at Lund University student perspectives Summary This report summarises the results of a survey that focused on the students experiences of writing a degree project at Lund University.
(From the Department of Anatomy, Harvard Medical School, Boston)
") THE FINE STRUCTURE OF THE ELECTRIC ORGAN OF THE ELECTRIC EEL AND TORPEDO RAY* PRELIMINARY COMMUNICATION BY JOHN H. LUFT, M.D. (From the Department of Anatomy, Harvard Medical School, Boston) PLATE 76 Electric
THE FINE STRUCTURE OF THE ELECTRIC ORGAN OF THE ELECTRIC EEL AND TORPEDO RAY* PRELIMINARY COMMUNICATION BY JOHN H. LUFT, M.D. (From the Department of Anatomy, Harvard Medical School, Boston) PLATE 76 Electric
Determination of g using a spring
 INTRODUCTION UNIVERSITY OF SURREY DEPARTMENT OF PHYSICS Level 1 Laboratory: Introduction Experiment Determination of g using a spring This experiment is designed to get you confident in using the quantitative
INTRODUCTION UNIVERSITY OF SURREY DEPARTMENT OF PHYSICS Level 1 Laboratory: Introduction Experiment Determination of g using a spring This experiment is designed to get you confident in using the quantitative
Spinal Anatomy. * MedX research contends that the lumbar region really starts at T-11, based upon the attributes of the vertebra.
 Spinal Anatomy Overview Neck and back pain, especially pain in the lower back, is one of the most common health problems in adults. Fortunately, most back and neck pain is temporary, resulting from short-term
Spinal Anatomy Overview Neck and back pain, especially pain in the lower back, is one of the most common health problems in adults. Fortunately, most back and neck pain is temporary, resulting from short-term
Golf Swing. Golf Swing Mechanics. Address Position. Posture To position the upper body correctly you must have the proper stance as described above.
 The golf swing is just that, a swing. In nearly all sports that involve swinging an implement, a bat, a hockey stick or racquet, the object to be struck is moving. In golf, the ball is always stationary.
The golf swing is just that, a swing. In nearly all sports that involve swinging an implement, a bat, a hockey stick or racquet, the object to be struck is moving. In golf, the ball is always stationary.
Victorian 'Live Box' Microscope
 Victorian 'Live Box' Microscope Capability in 40mm R. Jordan Kreindler (USA) Small cylindrical microscopes were popular at the end of the 19th century and the beginning of the 20th. Fig 1. presents some
Victorian 'Live Box' Microscope Capability in 40mm R. Jordan Kreindler (USA) Small cylindrical microscopes were popular at the end of the 19th century and the beginning of the 20th. Fig 1. presents some
6.4 Normal Distribution
 Contents 6.4 Normal Distribution....................... 381 6.4.1 Characteristics of the Normal Distribution....... 381 6.4.2 The Standardized Normal Distribution......... 385 6.4.3 Meaning of Areas under
Contents 6.4 Normal Distribution....................... 381 6.4.1 Characteristics of the Normal Distribution....... 381 6.4.2 The Standardized Normal Distribution......... 385 6.4.3 Meaning of Areas under
Evolution (18%) 11 Items Sample Test Prep Questions
 11 Items Sample Test Prep Questions") Evolution (18%) 11 Items Sample Test Prep Questions Grade 7 (Evolution) 3.a Students know both genetic variation and environmental factors are causes of evolution and diversity of organisms. (pg. 109 Science
Evolution (18%) 11 Items Sample Test Prep Questions Grade 7 (Evolution) 3.a Students know both genetic variation and environmental factors are causes of evolution and diversity of organisms. (pg. 109 Science
Classification of Malocclusion
 Classification of Malocclusion What s going on here? How would you describe this? Dr. Robert Gallois REFERENCE: Where Do We Begin? ESSENTIALS FOR ORTHODONTIC PRACTICE By Riolo and Avery Chapter 6 pages
Classification of Malocclusion What s going on here? How would you describe this? Dr. Robert Gallois REFERENCE: Where Do We Begin? ESSENTIALS FOR ORTHODONTIC PRACTICE By Riolo and Avery Chapter 6 pages
Transverse Sections of the Spinal Cord
 Transverse Sections of the Spinal Cord The spinal cord is perhaps the most simply arranged part of the CNS. Its basic structure, indicated in a schematic drawing of the eighth cervical segment (Figure
Transverse Sections of the Spinal Cord The spinal cord is perhaps the most simply arranged part of the CNS. Its basic structure, indicated in a schematic drawing of the eighth cervical segment (Figure
Attribution Guide. Variety 1 Small Head; straight line of curls at back of head. Large Head; interrupted line of curls at back of head
 Classic Head Quarter Eagles The Coinage Act of 1834 reduced the standard weight of gold coins to promote coinage and circulation of the coins, previously valued at more than face value. The specifications
Classic Head Quarter Eagles The Coinage Act of 1834 reduced the standard weight of gold coins to promote coinage and circulation of the coins, previously valued at more than face value. The specifications
SKELETON AND JOINTS G.C.S.E. PHYSICAL EDUCATION. Unit 1. Factors Affecting Participation and Performance. G.C.S.E. P.E. Teacher:.
 G.C.S.E. PHYSICAL EDUCATION Unit 1 Factors Affecting Participation and Performance SKELETON AND JOINTS Name: G.C.S.E. P.E. Teacher:. By the end of this booklet you should be able to: Understand what the
G.C.S.E. PHYSICAL EDUCATION Unit 1 Factors Affecting Participation and Performance SKELETON AND JOINTS Name: G.C.S.E. P.E. Teacher:. By the end of this booklet you should be able to: Understand what the
Biological kinds and the causal theory of reference
 Biological kinds and the causal theory of reference Ingo Brigandt Department of History and Philosophy of Science 1017 Cathedral of Learning University of Pittsburgh Pittsburgh, PA 15260 E-mail: inb1@pitt.edu
Biological kinds and the causal theory of reference Ingo Brigandt Department of History and Philosophy of Science 1017 Cathedral of Learning University of Pittsburgh Pittsburgh, PA 15260 E-mail: inb1@pitt.edu
CHA SERIES. Key Chiropractic Concepts for the CHA. Ontario Chiropractic Association. Treatment That Stands Up.
 CHA SERIES Key Chiropractic Concepts for the CHA AGENDA Welcome & Introductions About Chiropractic Terminology ABOUT CHIROPRACTIC You will get lots of questions about the profession & chiropractic care
CHA SERIES Key Chiropractic Concepts for the CHA AGENDA Welcome & Introductions About Chiropractic Terminology ABOUT CHIROPRACTIC You will get lots of questions about the profession & chiropractic care
Carnivore, omnivore or herbivore?
 Carnivore, omnivore or herbivore? Physical adaptations of the giant panda Student booklet (ST) October 2010 panda_st_student.doc Context The Giant Panda is a species that is faced with extinction. It is
Carnivore, omnivore or herbivore? Physical adaptations of the giant panda Student booklet (ST) October 2010 panda_st_student.doc Context The Giant Panda is a species that is faced with extinction. It is
climate science A SHORT GUIDE TO This is a short summary of a detailed discussion of climate change science.
 A SHORT GUIDE TO climate science This is a short summary of a detailed discussion of climate change science. For more information and to view the full report, visit royalsociety.org/policy/climate-change
A SHORT GUIDE TO climate science This is a short summary of a detailed discussion of climate change science. For more information and to view the full report, visit royalsociety.org/policy/climate-change
TMMC&RC, Teerthanker Mahaveer University, Moradabad, Uttar Pradesh, India, 244001 Mobile No.: +91-9997605146, E Mail drnidhivarshney@gmail.
 SEXUAL DIMORPHISM IN CERVICAL VERTEBRAL CANAL MEASUREMENTS OF HUMAN FOETUSES Nidhi Sharma *, Farah Ghaus, Nafis Ahmad Faruqi, Raghuveer Singh Mandloi * Correspondence to: Dr. Nidhi Sharma, Assistant Professor,
SEXUAL DIMORPHISM IN CERVICAL VERTEBRAL CANAL MEASUREMENTS OF HUMAN FOETUSES Nidhi Sharma *, Farah Ghaus, Nafis Ahmad Faruqi, Raghuveer Singh Mandloi * Correspondence to: Dr. Nidhi Sharma, Assistant Professor,
Making Tracks Elementary School 5-E Lesson
 Making Tracks Elementary School 5-E Lesson Objectives -Student will analyze fossil footprints of two and four legged dinosaurs to learn more about how these animals lived. -Students will experiment with
Making Tracks Elementary School 5-E Lesson Objectives -Student will analyze fossil footprints of two and four legged dinosaurs to learn more about how these animals lived. -Students will experiment with
Introduction. I. Objectives. II. Introduction. A. To become familiar with the terms of direction and location.
 E X E R C I S E Introduction I. Objectives A. To become familiar with the terms of direction and location. B. To become familiar with different types of planes and sections. C. To learn the names and locations
E X E R C I S E Introduction I. Objectives A. To become familiar with the terms of direction and location. B. To become familiar with different types of planes and sections. C. To learn the names and locations
July 2007. Management Practice & Productivity: Why they matter
 July 2007 Management Practice & Productivity: Why they matter Nick Bloom Stanford University Stephen Dorgan McKinsey & Company John Dowdy McKinsey & Company John Van Reenen Centre for Economic Performance,
July 2007 Management Practice & Productivity: Why they matter Nick Bloom Stanford University Stephen Dorgan McKinsey & Company John Dowdy McKinsey & Company John Van Reenen Centre for Economic Performance,
PROPORTIONS. The new Golden Rules in dentistry. History. Abstract
 M PROPORTIONS The new Golden Rules in dentistry Dr. Alain Méthot Abstract Since the beginning Cosmetic Dentistry has been using the principles of Golden Proportion (1: 0.618) as a guideline for smile design...
M PROPORTIONS The new Golden Rules in dentistry Dr. Alain Méthot Abstract Since the beginning Cosmetic Dentistry has been using the principles of Golden Proportion (1: 0.618) as a guideline for smile design...
Different wing in pitchers of the myrmecophagous species
 Different wing in pitchers of the myrmecophagous species Sarracenia minor and S. rubra Miloslav Studnička Liberec Botanic Gardens Purkyňova 630/1 CZ-460 01 Liberec Czech Republic botangarden@volny.cz Keywords:
Different wing in pitchers of the myrmecophagous species Sarracenia minor and S. rubra Miloslav Studnička Liberec Botanic Gardens Purkyňova 630/1 CZ-460 01 Liberec Czech Republic botangarden@volny.cz Keywords:
Yasuhiro Yamada a, *, Ken McClay b
 Journal of Structural Geology 25 (2003) 1331 1336 www.elsevier.com/locate/jsg Application of geometric models to inverted listric fault systems in sandbox experiments. Paper 2: insights for possible along
Journal of Structural Geology 25 (2003) 1331 1336 www.elsevier.com/locate/jsg Application of geometric models to inverted listric fault systems in sandbox experiments. Paper 2: insights for possible along
Create a Poster Using Publisher
 Contents 1. Introduction 1. Starting Publisher 2. Create a Poster Template 5. Aligning your images and text 7. Apply a background 12. Add text to your poster 14. Add pictures to your poster 17. Add graphs
Contents 1. Introduction 1. Starting Publisher 2. Create a Poster Template 5. Aligning your images and text 7. Apply a background 12. Add text to your poster 14. Add pictures to your poster 17. Add graphs
Plate Tectonics: Ridges, Transform Faults and Subduction Zones
 Plate Tectonics: Ridges, Transform Faults and Subduction Zones Goals of this exercise: 1. review the major physiographic features of the ocean basins 2. investigate the creation of oceanic crust at mid-ocean
Plate Tectonics: Ridges, Transform Faults and Subduction Zones Goals of this exercise: 1. review the major physiographic features of the ocean basins 2. investigate the creation of oceanic crust at mid-ocean
What is a Fossil? Adapted from What is a Fossil MOR Dinosaur Trunk Activity
 What is a Fossil? Adapted from What is a Fossil MOR Dinosaur Trunk Activity Grade Level: Grades K-8 Preparation Time: 10-15 minutes Activity Duration: 30 minutes-2 hours Concepts Addressed A fossil is
What is a Fossil? Adapted from What is a Fossil MOR Dinosaur Trunk Activity Grade Level: Grades K-8 Preparation Time: 10-15 minutes Activity Duration: 30 minutes-2 hours Concepts Addressed A fossil is
How to increase Bat Speed & Bat Quickness / Acceleration
 How to increase Bat Speed & Bat Quickness / Acceleration What is Bat Speed? Bat Speed: Bat speed is measured in miles per hour (MPH) and considers only the highest speed of the bat head (peak velocity)
How to increase Bat Speed & Bat Quickness / Acceleration What is Bat Speed? Bat Speed: Bat speed is measured in miles per hour (MPH) and considers only the highest speed of the bat head (peak velocity)
Chapter 9 The Hip Joint and Pelvic Girdle
 Copyright The McGraw-Hill Companies, Inc. Reprinted by permission. The Hip Joint and Pelvic Girdle Chapter 9 The Hip Joint and Pelvic Girdle Structural Kinesiology R.T. Floyd, Ed.D, ATC, CSCS Hip joint
Copyright The McGraw-Hill Companies, Inc. Reprinted by permission. The Hip Joint and Pelvic Girdle Chapter 9 The Hip Joint and Pelvic Girdle Structural Kinesiology R.T. Floyd, Ed.D, ATC, CSCS Hip joint
Background Biology and Biochemistry Notes A
 Background Biology and Biochemistry Notes A Vocabulary dependent variable evidence experiment hypothesis independent variable model observation prediction science scientific investigation scientific law
Background Biology and Biochemistry Notes A Vocabulary dependent variable evidence experiment hypothesis independent variable model observation prediction science scientific investigation scientific law
THE SKELETAL SYSTEM FUNCTIONS OF THE SKELETAL SYSTEM
 THE SKELETAL SYSTEM The skeleton is the body s bony framework which consists of 206 bones. The bones are made up of water(45%), calcium and phosphorous(35%) and other organic materials(20%). The calcium
THE SKELETAL SYSTEM The skeleton is the body s bony framework which consists of 206 bones. The bones are made up of water(45%), calcium and phosphorous(35%) and other organic materials(20%). The calcium
Unit 7: Normal Curves
 Unit 7: Normal Curves Summary of Video Histograms of completely unrelated data often exhibit similar shapes. To focus on the overall shape of a distribution and to avoid being distracted by the irregularities
Unit 7: Normal Curves Summary of Video Histograms of completely unrelated data often exhibit similar shapes. To focus on the overall shape of a distribution and to avoid being distracted by the irregularities
Chapter 11. What are the functions of the skeletal system? More detail on bone
 Skeletal System Chapter 11 11.1 Overview of the skeletal system What are the functions of the skeletal system? 1. Supports the body 2. Protects the soft body parts 3. Produces blood cells 4. Stores minerals
Skeletal System Chapter 11 11.1 Overview of the skeletal system What are the functions of the skeletal system? 1. Supports the body 2. Protects the soft body parts 3. Produces blood cells 4. Stores minerals
MD5-26 Stacking Blocks Pages 115 116
 MD5-26 Stacking Blocks Pages 115 116 STANDARDS 5.MD.C.4 Goals Students will find the number of cubes in a rectangular stack and develop the formula length width height for the number of cubes in a stack.
MD5-26 Stacking Blocks Pages 115 116 STANDARDS 5.MD.C.4 Goals Students will find the number of cubes in a rectangular stack and develop the formula length width height for the number of cubes in a stack.
CHAPTER 10 RESTS AND PREPARATIONS. 4. Serve as a reference point for evaluating the fit of the framework to the teeth.
 CHAPTER 10 RESTS AND DEFINITIONS A REST is any rigid part of an RPD framework which contacts a properly prepared surface of a tooth. A REST PREPARATION or REST SEAT is any portion of a tooth or restoration
CHAPTER 10 RESTS AND DEFINITIONS A REST is any rigid part of an RPD framework which contacts a properly prepared surface of a tooth. A REST PREPARATION or REST SEAT is any portion of a tooth or restoration
LUMBAR LAMINECTOMY AND DISCECTOMY. Basic Anatomical Landmarks: Posterior View Lumbar Spine
 Lumbar Relating to the loins or the section of the back and sides between the ribs and the pelvis. In the spinal column, the last five vertebrae (from superior to inferior, L1-L5) Laminectomy Surgical
Lumbar Relating to the loins or the section of the back and sides between the ribs and the pelvis. In the spinal column, the last five vertebrae (from superior to inferior, L1-L5) Laminectomy Surgical